










JMLS 2023 June;8(1):1-7. 10.23005/ksmls.2023.8.1.1 Epub 2023 June 16
Copyright © 2023 by The Korean Society of Marine Life Science
Microanatomical Structure of Male Reproductive Organ in the Common Squid, Todarodes pacificus (Cephalopoda: Ommastrephidae)
So Ryung Shin; Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
Jung Sick Lee; Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
- Abstract
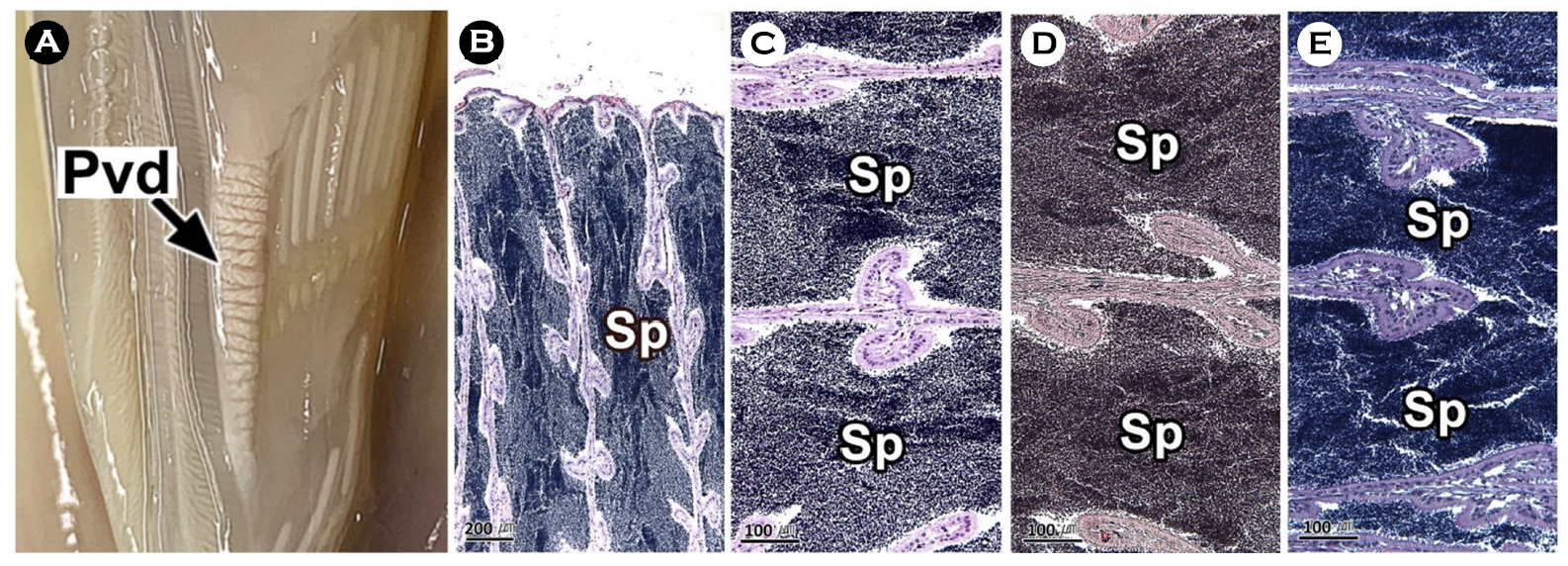
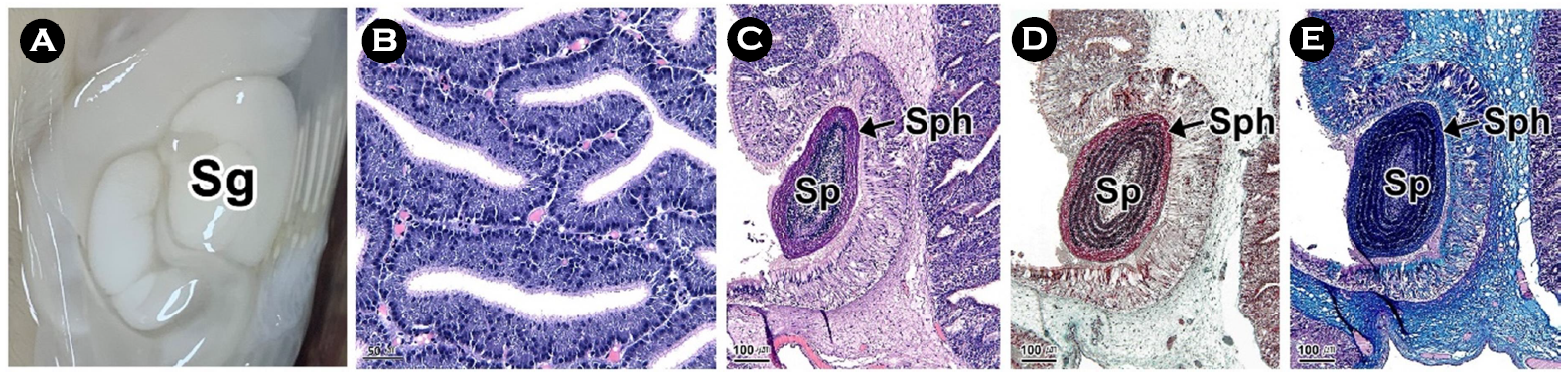
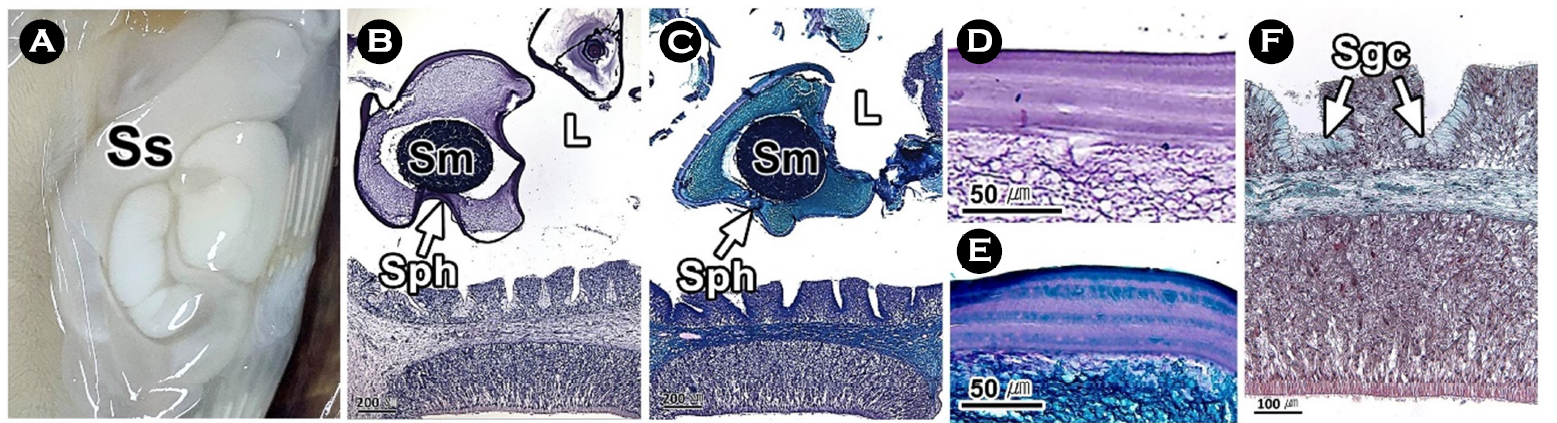
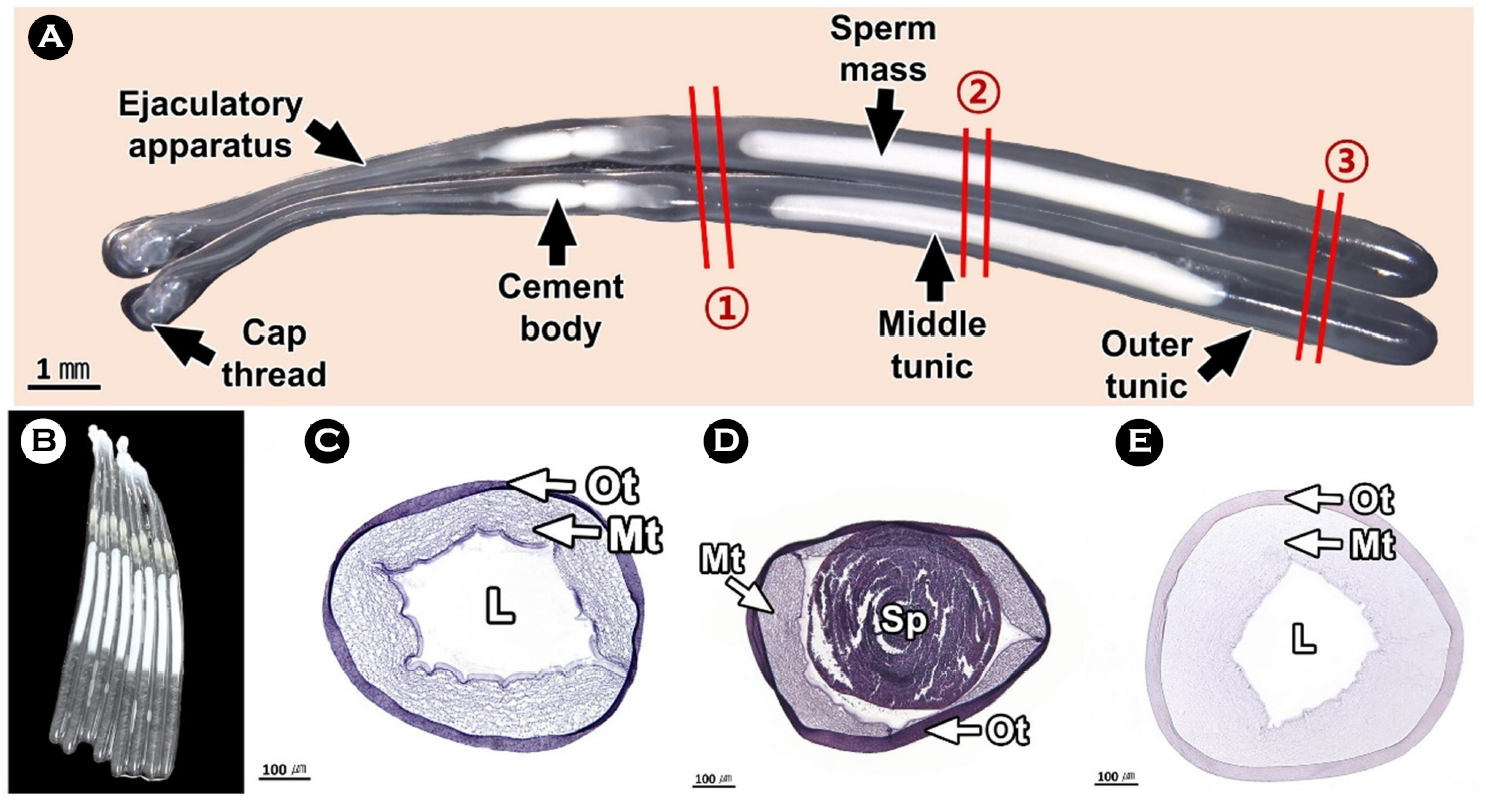
연구에서는 두족류의 생식생태를 이해하기 위하여 우리나라 주요 두족류인 살오징어 Todarodes pacificus 수컷의 생식기관과 정포의 미세해부학적 구조를 기재하였다. 살오징어는 자웅이체로 외투막에 투과되는 생식기관의 색과 교접완을 통하여 암, 수를 구분할 수 있는 성적이형을 가진다. 수컷 생식기관은 정소, 1차 수정관, 정포선, 정포낭, 2차 수정관으로 구성된다. 살오징어의 수컷은 정포 형성기관인 정포선과 같은 특수화된 생식기관을 가진다. 정소는 조직학적으로 정세관형이었다. 1차 수정관은 정소의 후방부터 정포선까지 연결된 가늘고 긴 역삼각형의 용수철 형태였다. 내부에는 H-E 염색에서 호염기성으로 반응하는 정자가 가득 차 있었다. 정포선은 1차 수정관과 정포낭에 연결된 불규칙한 타원형으로 내부에는 다수의 관상선이 존재하였다. 정포낭은 정포선과 2차 수정관 사이에 위치하는 관 구조로 내강에서는 다수의 정포가 확인되었다. 2차 수정관은 정포낭의 후방부에 연결되며, 내부에 정포를 갖고 있었다. 정포는 길이 약 22 mm의 길고 투명한 관 형태로 내부에는 정자 덩어리를 갖고 있었다.
This study was performed to understand the reproductive ecology of cephalopods, described the microanatomical structure of the male reproductive organs and spermatophore in the common squid, Todarodes pacificus, a major cephalopods in Korea. The common squid was gonochorism and had sexual dimorphism, the color of the reproductive organs reflected on the mantle and the presence of the hectocotylus. Male reproductive organs were composed of testis, primary vas deferens, spermatophoric gland (seminal vesicle), spermatophoric sac (Needham's sac) and secondary vas deferens. The male has specialized reproductive organs such as the spermatophore forming organ, spermatophoric gland. Testis was histologically a seminiferous tubule type. The primary vas deferens was a thin and inverted triangular spring-like form that connected from the rear of the testis to the spermatophoric gland. Inside, it was filled with sperm of basophilic in H-E stain. The spermatophoric gland is an irregular oval connected to the primary vas deferens and spermatophoric sac, and there were a number of tubular glands. The spermatophoric sac is a tubular structure located between spermatophoric gland and secondary vas deferens, and a number of spermatophores have been identified in the lumen. The secondary vas deferens was connected to the posterior of the spermatophoric sac and had a spermatophore inside. The spermatophore was a long, transparent tube about 22 mm long, with a sperm mass inside.
Keywords: Todarodes pacificus(살오징어) Male reproductive organ(수컷 생식기관) Spermatophore(정포)
Correspondence to: Jung Sick Lee; Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
- Received
- 29 May 2023;
- Revised
- 31 May 2023;
- Accepted
- 3 June 2023.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
두족류는 성장이 매우 빠르고 일생에 한번의 생식과정을 거친 후 죽는 것으로 알려져 있다(Mangold and Boyle, 1987). 이와 같이 수명이 짧은 종의 생식활동은 물리적, 생물학적 환경에 크게 좌우된다(Lipiński et al., 1998). 수서동물의 수정은 수정장소에 따라 체외수정과 체내수정으로 구분되는데, 체내수정종은 일반적으로 정자를 현탁액의 형태로 암컷에게 전달한다. 하지만 도롱뇽 Hynobius leechii (Dunn, 1923)을 비롯하여 경골어류인 인상어 Neoditrema ransonneti (Lee et al., 2001), 갑각류(Subramoniam, 1993), 두족류(Marian, 2015) 수컷은 정자를 정포(spermatophore) 형태로 암컷에게 전달한다. 정포를 갖는 종은 정포 형성을 위한 분비선과 같은 생식기관이 분화되어 있다(Drew, 1919; Lopez Greco et al., 2007). 두족류 가운데 Loligo pealii에서 정포는 정소에서 만들어진 정자가 수정관을 통해 정포선으로 이동하면서 형성된다(Drew, 1919). 하지만 살오징어 Todarodes pacificus에서 이러한 정포 형성기관에 관한 연구는 전무하다.
살오징어는 개안목(Oegopsida), 살오징어과(Ommastrephidae)에 속하는 두족류로 한국에서는 참살오징어속(Todarodes)에 속하는 유일한 종으로 한국을 비롯한 일본, 중국에서 상업적으로 매우 중요한 위치에 있다(Worms, 2023). 본 연구는 살오징어 수컷의 생식기관과 정포의 구조를 기재하여 생식생물학적 기초자료를 제공하고자 하였다.
재료 및 방법
1. 재료
본 연구에서는 살오징어 성체(ML 19.3 cm)의 생체시료(n=20)를 사용하였다(Fig. 1).

2. 방법
2.1. 해부학적 구조
살오징어의 해부학적 구조는 해부 후, 외투강 내 기관계의 위치와 형태 등을 이미지 촬영 및 해부현미경을 이용하여 기재하였다.
2.2. 조직학적 분석
광학현미경 표본은 생식기관을 적출하여 2.5% glutaraldehyde 용액에 24시간 고정한 후, 파라핀 절편법에 따라 3~5 μm 두께로 연속절편하여 제작하였다. 조직표본은 Mayer's hematoxylin-0.5% eosin (H-E) 이중염색, Masson 삼중염색, alcian blue-periodic acid and Schiff’s solution (AB-PAS, pH 2.5) 반응을 실시하였다.




- References
-
1. Anadón R. 2019. Functional histology: the tissues of common coleoid cephalopods. Gestal C, Pascual S, Guerra Á, Fiorito G, Vieites JM (ed.), Handbook of pathogens and diseases in cephalopods. Springer, Berlin, pp 39-86.

-
2. Arnold JM, Williams-Arnold LD. 1977. Cephalopoda: Decapoda. Giese AC, Pearse JS (ed.), Reproduction of marine inver- tebrates. Academic press, London, pp 243-290.
-
3. Austin CR, Mann CL, Mann TRR. 1964. Spermatophores and spermatozoa of the squid Loligo pealii. Proc R Soc Lond B 161: 143-152.
-
4. Blancquaert T. 1925. L'origine et la formation des spermatophores chez les céphalopodes décapodes. Cellule 36: 315-356.
-
5. Cuccu D, Mereu M, Agus B, Cau A, Culurgioni J, Sabatini A, Jereb P. 2014. Male reproductive system and spermatophores pro- duction and storage in Histioteuthis bonnellii (Cephalopoda: Histioteuthidae): a look into deep-sea squids' reproductive strategy. Deep Sea Res Part I: Oceanogr 91: 86-93.
-
6. Drew GA. 1919. Sexual activities of the squid Loligo pealii (Les.). II. The spermatophore; its structure, ejaculation and formation. J Morphol 32: 379-435.
-
-
8. Hanlon RT, Messenger JB. 2018. Reproductive behavior. Hanlon RT, Messenger JB (ed.), Cephalopod behaviour. Cambridge University Press, Cambridge, pp 148-205.
-
9. Lee JS, Jeong SY, Chung EY. 2001. Formation and structure of the spermatozeugmata of Neoditram ransonneti (Perciformes: Embiotocidae). Dev Reprod 5: 145-150.
-
10. León-Guzmán SS, Alejo-Plata MDC, Morales-Bojórquez E, Benítez-Villalobos F. 2020. Reproductive biology of the dart squid, Lolliguncula diomedeae (Cephalopoda: Loliginidae) from Gulf of Tehuantepec, Mexico. Mar Biol Res 16: 327-339.
-
11. Lipiński MR, Butterworth, DS, Augustyn CJ, Brodziak JKT, Christy G, des Clers S, Jackson GD, O'Dor RK, Pauly D, Purchase LV, Roberts MJ, Roel BA, Sakurai Y, Sauer WHH. 1998. Cepha- lopod fisheries: a future global upside to past overexploitaion of living marine resources? Results of an International Work- shop, 31 August - 2 September 1997, Cape Town, South Africa. S Afr J Mar Sci 20: 463-469.
-
12. Lopez Greco LS, Vazquez F, Rodríguez EM. 2007. Morphology of the male reproductive system and spermatophore formation in the freshwater 'red claw' crayfish Cherax quadricarinatus (Von Martens, 1898) (Decapoda, Parastacidae). Acta Zool 88: 223-229.
-
13. Mangold K, Boyle PR. 1987. Reproduction. Boyle PR (ed.), Cepha- lopod life cycles. Academic Press, London, pp 157-200.
-
14. Mann T, Martin AW, Thiersch JB. 1970. Male reproductive tract, spermatophores and spermatophoric reaction in the giant octopus of the North Pacific, Octopus dofleini martini. Proc R Soc Lond B 175: 31-61.
-
15. Marian JEAR. 2015. Evolution of spermatophore transfer mech- anisms in cephalopods. J Nat Hist 49: 1-33.
-
16. Nigmatullin CM, Sabirov RM, Zalygalin VP. 2003. Ontogenetic aspects of morphology, size, structure and production of spermatophores in ommastrephid squids: an overview. Berliner Paläobiol Abh 3: 225-240.
-
17. Sato N, Yoshida MA, Fujiwara E, Kasugai T. 2013. High-speed camera observations of copulatory behaviour in Idiosepius paradoxus: function of the dimorphic hectocotyli. J Molluscan Stud 79: 183-186.
-
18. Sauer WH, Lipiński MR. 1990. Histological validation of morpho- logical stages of sexual maturity in chokker squid Loligo vulgaris reynaudii D'Orb (Cephalopoda: Loliginidae). S Afri J Mar Sci 9: 189-200.
-
19. Sakurai Y, Bower JR, Ikeda Y. 2003. Reproductive characteristics of the ommastrephid squid Todarodes pacificus. Kjesbu OS, Hunter JR, Witthames PR (ed.), Report of the working group on modern approaches to assess maturity and fecundity of warm-and cold-water fish and squids. Havforskningsinstituttet, Bergen, pp 105-116.
-
20. Subramoniam T. 1993. Spermatophores and sperm transfer in marine crustaceans. Adv Mar Biol 29: 129-214.
-
21. Worms, World Resister of Marine Species. 2023. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=342067 (accessed on 8 Jun. 2023).
Copyright (C) 2025. The Korean Society of Marine Life Science