










JMLS 2017 October;2(2):83-89. Epub 2017 November 03
Copyright © 2017 by The Korean Society of Marine Life Science
Molecular Identification and Effects of Temperature on Survival and Growth of Hybrids between Haliotis gigantea Gmelin (♀) and Haliotis discus hannai Reeve (♂)
Hye Suck An; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Jong Won Han; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Hyun-Ju Hwang; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Hancheol Jeon; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Seung-Hyun Jung; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Seonmi Jo; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Tae-Young Choi; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Young Se Hyun; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Ha Yeun Song; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
Ilson Whang; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea
- Abstract
In abalones, interspecific hybridization has been suggested as a possible means to increase production and desired traits for the industry. In Korea, Haliotis gigantea is considered a species with a larger size and higher temperature tolerance than H. discus hannai. However, H. discus hannai is considered the most valuable and popular fishery resource due to its better acceptance and higher market prices. Thus, viable interspecific hybrids have been produced by artificial inseminating H. gigantea eggs with H. discus hannai sperm. However, the reciprocal hybrid cross was not successful. In this study, the hybridity and the growth and thermal tolerance performance of the interspecific hybrids were examined. A combination of various assays revealed maximum growth occurrence at 21℃ and the higher growth rate in the hybrids than that of H. discus hannai parent. In addition, the growth and survival at high-temperature (28℃) of the hybrids was equivalent to that of the highly tolerant H. gigantea parent, suggesting new possibilities to overcome the mass mortality in H. discus hannai during high temperature periods of summer season in Korea. Furthermore, the induced interspecific hybrid status was confirmed by the presence of species-specific bands for each parental species of the random amplified polymorphic DNA (RAPD) profiles using universal rice primer (URP), which could be used as species-specific markers to distinguish the hybrids and their parental species.
Correspondence to: Hye Suck An; Genetic Resources Research Department, National Marine Biodiversity Institute of Korea (MABIK), Chungchungnam-do 33662, Korea; E-mail : mgran@mabik.re.kr;
- Received
- 27 September 2017;
- Revised
- 20 October 2017;
- Accepted
- 26 October 2017.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: English,
Full Text:

Introduction
Abalone are a group of marine gastropod mollusks belonging to the genus Haliotis that comprises over 100 species and are distributed throughout of the world in temperate and tropical oceans (Geiger and Poppe, 2000). Among the more than 30 species of commercially available abalone, six species of Pacific abalone have been reported to occur along the Korean coasts, H. discus hannai Ino, H. discus discus Reeve, H. madaka Habe, H. gigantea Gmelin, H. diversicolor supertexta Reeve, and H. diversicolor diversicolor Reeve (Kim et al., 1988; An et al., 2005). Of these, H. discus hannai is the most commercially valuable species and a highly sought-after delicacy in Korea. Seedling production and rearing techniques for H. discus hannai have been established and its aquaculture has become increasingly popular because of its consistent high demand and market value (Zhang et al., 2004). The production of this abalone increased drastically from about 1,000 t/y in 2003, reaching 6,941 t in 2011 (wild catch, 162 t and aquaculture, 6,779 t; Korean Statistical Office, 2012). H. gigantea is a valued commercial species distributed along the coast of Jeju Island, Korea. However, the production has not reached important levels due to the poor performance. This species is considered a species with a larger size and higher temperature tolerance than H. discus hannai. The meat of H. gigantea is crisp and tender, and this species possesses excellent disease resistance (Gao et al., 2000). These traits make it a commercial aquaculture abalone species in Korea. H. discus hannai and H. gigantea are easily distinguished by their surface features and by the color of their shells.
Currently, the production of intra- and interspecific hybrids is an alternative to conventional selective breeding methods of economically important aquaculture species (Elliott, 2000). Hybridization has been successfully obtained in many marine species (Bartley et al., 2001; Hulata, 2001). In worldwide abalone aquaculture, more than 50 crosses between sympatric and/or allopatric species have been reported to be successfully produced and most of the hybrid ablone evaluated have shown faster growth, high survival rate, particular adaptations to cultured conditions and desirable market qualities (Lafarga-de la Cruz and Gallardo-Escárate, 2010). Thus, hybridization between H. discus hannai and H. gigantea has been suggested as a possible method to increase growth rate and adaptability to environmental conditions. Laboratory crosses of H. discus discus and H. gigantea have demonstrated that F1 hybrids can be produced with a low fertilization rate (An et al., 2007; Ahmed et al., 2008). Genetic identification and biological characterization of hybridized individuals are the first step for developing conservation and management strategies and improving the aquaculture.
Advances in molecular genetics in recent years have provided several types of genetic markers to be used to identify inter- and intraspecies levels and hybridization in aquatic animals (Liu and Cordes, 2004; Choi et al., 2012). Among them, the random amplified polymorphic DNA-polymerase chain reaction (RAPD-PCR) technique is a relatively simple and cheap method capable of differentiating taxa without the need to know their genomes (Williams et al., 1990). RAPD markers have been used to study species identification and genetic relationships for many other marine organisms including abalone (Klinbunga et al., 2004). URPs (universal rice primers) were developed from repetitive sequence of rice genome (Kang et al., 2002). URP-PCR technique is a useful tool for the characterization and grouping most eukaryotic or prokaryotic genomes, especially at inter- and intraspecies levels (Kang et al., 2002; Jana et al., 2005).
Temperature is a direct and primary environmental factor determining an aquatic organism's activity and therefore mobile species including the abalones show different behavioral responses. Hence, when culturing a species, best performance is obtained when temperatures are optimal (Díaz et al., 2000). Furthermore, high temperature during summer is apparently a major constraint for abalone farming in Korea. Therefore, the thermal threat has become one of the major challenges for abalone farming in Korea. An efficient and moderate method that can precisely predict abalone thermal performance would be greatly beneficial in breeding projects. Thus, ascertaining effects of temperature on survival and growth of the hybrids is very important due its pivotal role for the abalone physiology.
The main objective of this study was to confirm the hybridity of the hybrids between H. gigantea (♀) and H. discus hannai (♂) using URP-PCR approach and to evaluate effects of temperature on growth and survival of the hybrids in order to assess the ability to adapt to different thermal regimes. These data will be useful for developing conservation and management strategies and improving the aquaculture industry of abalone fisheries in Korea.
Materials and Methods
1. Samples
The broodstock of H. gigantea (G) and H. discus hannai (D) were originally from Jeju Island and Pohang, Korea respectively. Two species were collected between February and May of 1999 and 2000.
Hybrids between H. gigantea and H. discus hannai was produced (GD, H. gigantea ♀ × H. discus hannai ♂) and two purebred controls (DD, H. discus hannai ♀ × H. discus hannai ♂ and GG, H. gigantea ♀ × H. gigantea ♂) in 2000 at the Shellfish Genetic and Breeding Research Center, Jeju Fisheries Research Institute, National Fisheries Research and Development Institute, Jeju Island, Korea. After artificial insemination, the mantle muscle of parental abalones were sampled and stored in 99% ethanol for DNA ex- traction. The hybrids and progeny of purebred were reared at tanks separately to prevent intermix. Young abalones from each genotype were raised in multiple tanks per genotype and fed microalgae that grew on the surfaces of the collectors until their shell length was over 6 mm. After that, they were kept in a cage made from netting with a mesh opening size of 2 mm and fed marine algae.
For molecular identification of the hybrid, parental abalones and live juvenile abalones of GD cross, which were randomly sampled at four months of age, were used for RAPD analysis.
For growth and thermal tolerance analysis, live juvenile abalones of each cross were sampled at seven months of age (approximately 1.05 cm shell length and 0.13 g body weight) and reared at each temperature.
2. DNA extraction
Total genomic DNA was extracted from mantle musculature samples of the juvenile abalone of GD cross and parental abalones. The removed musculatures were minced and lysed by TNES-Urea buffer (6 or 8 M urea: 10 mM Tris- HCL, pH 7.5: 125 mM NaCl: 10 mM EDTA: 1% SDS) (Asahida et al., 1996). The genomic DNA was precipitated from the lysate with 100% ethanol. Following an ethanol wash the DNA was solubilized in water (or 8 mM NaOH). DNA was diluted with water to a final concentration of 20 ng/μl.
3. Genetic identification of the hybridity
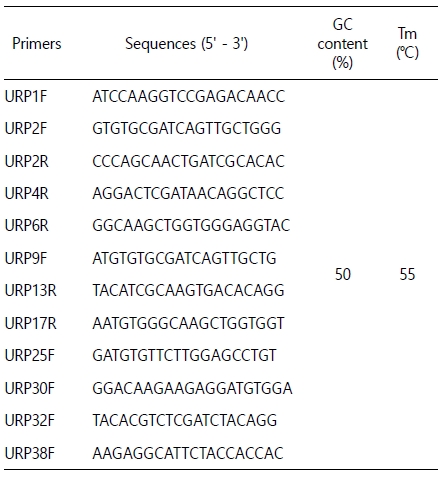
Primers and PCR conditions are adopted from Kang et al. (1997). Sequences of URP primers are listed in Table 1. Amplification of abalone genomic DNA using URP primers was performed in a PTC-100 thermal cycler (MJ Research, USA). PCR reactions were done in 50 μl volumes. Each reaction contained 5 μl of 10X EX TaqTM buffer (Takara, Shiga, Japan), 0.2 mM each dNTP, 2 μl of each primer (100 ng), 2.5 units of EX TaqTM polymerase (Takara), and 5 μl genomic DNA. The temperature regime used for the amplification was 35 repetitions of a three-step cycle consisting of denaturation at 95℃ for 1 min, annealing at 55℃ for 1 min, and extension at 72℃ for 2 min. Denaturation for the first cycle was 4 min at 95℃ and extension for the final cycle was 7 min at 72℃. Fifteen μl URP-PCR products were electrophoresed in 2% agarose gel with TAE buffer. DNA fragments in the gel were visualized by staining with ethidium bromide and photographed under UV transilluminator.
The primers that produced good amplification (clearly defined bands that differentiated between parental abalones) were used in the next amplification step. At this stage, primers were considered as informative and reliable for identification of the hybrid when they produced a specific amplicon both in all individuals of each parental abalone and in all individuals of hybrid.
4. Effects of temperature on survival and growth of the hybrids
In order to investigate effects of temperature on growth and survival of the hybrid and their purebred abalones, the experiments carried out indoors in a flow through system consisting of 30 tanks (35 cm in width × 28.5 cm in length × 23 cm in height) that each served as water baths. A thermoregulator was connected to one extreme for cooling and a 1,000 W heater was placed in the other extreme to keep a thermal temperature between 7~ 28℃. The vertical thermal stratification was eliminated by use of air stones placed in each tank.
The experiment had two replicate designs with five temperatures [ambient (16.7~28.1℃), 7, 14, 21, and 28℃] per three crosses (GD, DD and GG). Overall 30 trials were conducted. Each trial, 30 juvenile abalones of each cross (mean 1.05 cm initial shell length and mean 0.13 g weight) were allocated to each tank. Abalones were exposed to each temperature for 136 days (14-May, 2001~ 27-Sep, 2001). Feed was offered twice a week. All treatments were offered the same amount of feed as the tanks that had the largest biomass, thus ensuring that ration size did not further increase the growth of faster growing abalone. Mortalities were checked daily during the experiments and recorded and removed. Every month after the start of the experiments, individual weight and length of all abalones from each tank were measured.
Data were analyzed using SYSTAT 10 statistics software. Differences in survival and growth of each phenotype were assessed by ANOVA (followed by Duncan's multiple range test). Significance was set at p < 0.05.
Results
1. Genetic evidence for hybridisation
The RAPD-PCR technique can produce non-reproducible amplification products (Harry et al., 1998). Thus, reactions were performed following strict protocol with standardized conditions. Also, all amplification reactions were carried out, at least, three times in order to score reproducible bands clearly.
Hybrid identification was performed by 12 different URP primers (Table 1). They were used as template to assess the wide distribution of the URP nucleotide motif sequences in the parental abalone species genome. The number of amplified bands detected varied, depending on the primers, parental species and individuals. Out of these 12 primers screened, five primers generated reproducible profiles (URP1F, URP2F, URP2R, URP6R and URP30F) and showed different profiles between two parental species. The rest seven primers did not amplify at all or produced highly inconsistent amplification products from the same individuals and were excluded from further analysis. Among 5 primers URP2R only produced both unambiguous patterns of their parents in the hybrid. The gel pictures showing the banding patterns of RAPD profiles generated by URP2R are shown in Fig. 1. The male H. discus hannai had two specific amplicons B1 and B2 only. The female H. gigantea had a specific amplicon B3 only. The hybrid had a heterotic combination of all parental species specific amplicons B1, B2 and B3.
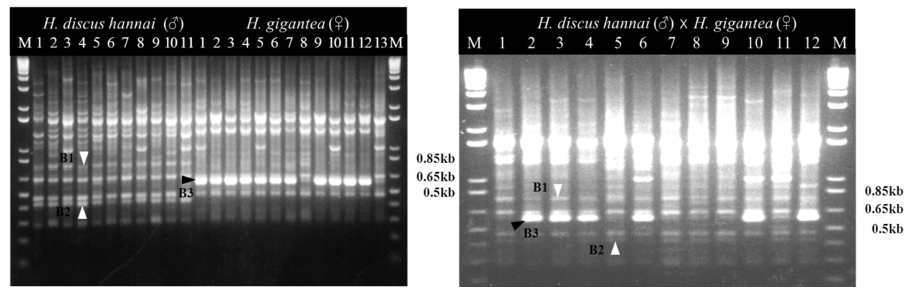
2. Effects of temperature on survival and growth of hybrids
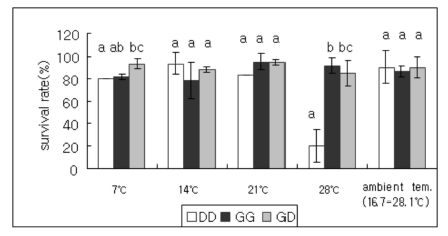
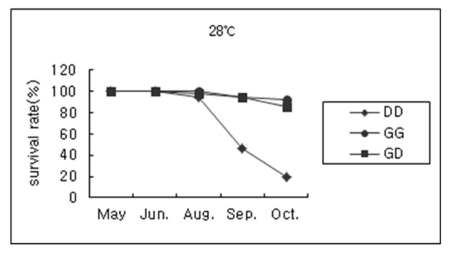
In order to examine the effect of temperature on survival of hybrids, observations on the survival rate of juvenile abalones of DD, GG and GD crosses were studied at ambient (16.7~28.1℃), 7, 14, 21, and 28℃, respectively. Overall, optimal culturing temperatures of ambient, 14 or 21℃ had no effect on survival of juvenile abalones of three crosses. However, the survivals at low and high-temperature (7 and 28℃) of the hybrids were equivalent to that of the highly tolerant H. gigantea parent (Fig. 2). This result may reflect a heterotic effect on survival at low and high water temperatures in the case of the hybrid between the two species compared with those of juvenile abalones of DD cross. Especially, the survival rate of juvenile abalones of DD cross cultured at 28℃ was rapidly reduced after 3 months exposure (Fig. 3).
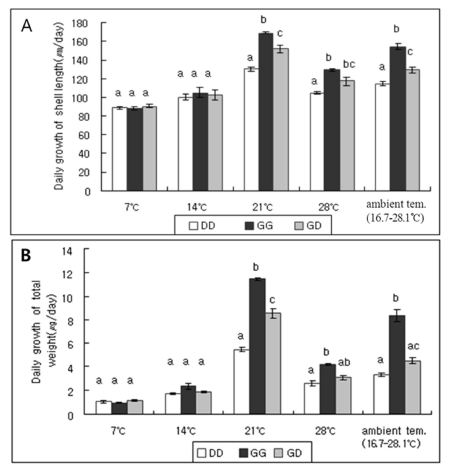
In order to examine the effect of temperature on the growth characteristics of the hybrid, observations on the growth rate of juvenile abalones of DD, GG and GD crosses were studied at ambient (16.7~28.1℃), 7, 14, 21, and 28℃, respectively. Fig. 4 shows the daily growth in shell length and total weight for juvenile abalones of three crosses. Overall, low culturing temperatures of 7 and 14℃ had no effect on the growth rate of juvenile abalones of three crosses. However, at ambient, 21 and 28℃, the values for juvenile abalones of GG cross (154.23±3.26, 169.11±1.43 and 129.22±0.83, respectively) were followed by those for the hybrid (129.61±3.59, 152.25±3.93 and 117.42±4.48, respectively), which was significantly higher than those of juvenile abalones of DD cross (114.70±2.02, 130.03±2.21 and 104.79±1.62, respectively) (p < 0.05). Therefore, the growth rates of the hybrid were intermediate between parental species at these relatively high temperatures (Fig. 4, A). Similar tendency was also detected in the daily growth in total weight for juvenile abalones of three crosses except no significant difference between the hybrid and juvenile abalones of DD cross at ambient and 28℃ (Fig. 4, B).
Discussion
Abalone hybrid production is difficult and success may reflect genetic affinities between species. A variety of approaches have been used to identify natural or artificial induced hybrids. The verification includes the use of morphological evidences, allozyme and RAPD markers. However, morphological differences of hybrids such as shell characteristics are only minor and need close examination by experienced workers and it is difficult to verify the genuine hybrids from interspecific hybridization (Ibarra et al., 2005). Previous studies using allozymes indeed described the genetic difference between the hybrids and their parental species (Ibarra et al., 2005; Ahmed et al., 2008). However, the use of allozyme loci failed to provide a sufficient genetic basis of hybrids, probably due to the less informative nature of allozyme loci. Mutation at the DNA level that causes a replacement of a similarly charged amino acid may not be detected by allozyme electrophoresis, although allozymes represent actual gene products. The information provided by dominant markers yielded by RAPD primers could be useful to facilitate the detection of genetic contributions of different species (Rajora and Rahman, 2003). The main reason for the success of RAPD analysis is the gain of a large number of genetic markers that require small amounts of DNA without the need of the taxa under study. There are many species in which such markers were used, including various abalones (Klinbunga et al., 2004; Marín et al., 2007).
Verification of the hybrid nature in interspecific crossing is indispensable for genetic improvement of abalones. In this study, we present reliable and highly reproducible procedures for the identification of hybrids between the male H. discus hannai and the female H. gigantea. H. discus hannai and H. gigantea are genetically very distinct. In our study, RAPD band profiles of URP2R primer allowed us to clearly distinguish both the parental species and their hybrids. As RAPD is the dominant marker, it has additional advantages with high sensitivity and high reproducibility. RAPD band profiles of the hybrids resulted in the presence of parent-specific bands, which were inherited from both parents. The information yielded by these bands could be sufficient for the identification of interspecific hybrids. Thus, the fact that the hybrid showed the simultaneous presence of the species-specific bands of both parental species indicates that genetic hybridization was accomplished in the cross between the male H. discus hannai and the female H. gigantea.
The pivotal role of temperature in controlling the rate of metabolic processes is well established and is reflected in survival and growth rates of poikilothermic animals including abalone. Several abalone hybrids have shown positive heterosis (hybrid vigour), such as faster growth and higher survival rates than at least one of the parental species (Lafarga-d la Cruz and Gallardo-Escárate, 2011). Our results concur with previous studies, showing that the survival and growth rates in shell length and total weight of juvenile abalone of H. gigantea were superior to those of H. discus hannai at water temperature 28℃ and the survival and growth rates of the hybrid was equivalent to those of the highly tolerant H. gigantea parent at water temperature 7 and above 21℃. Similarly, the juvenile hybrid between a female H. discus hannai and a male H. gigantea also showed growth superior to that of its parental species (Hara, 1992). Furthermore, our result also showed that the survival rate of juvenile abalones of H. discus hannai parent was rapidly decreased at high-temperature (28℃). Increasing water temperature can also act as a stressor impacting the immune responses of mollusks, associated with increased susceptibility to disease out-breaks (Dang et al., 2012). Therefore, temperature in- crease can be a major environmental stressor on marine organisms. When Pacific abalone was exposed to 28℃ for 7 days, a dramatic reduction in HSP70 expression occurred. This drop in HSP70 level of juvenile abalones of H. discus hannai might be caused by intolerance to long-term exposure at high temperature and this intolerance had caused the breakdown of homeostasis in these juvenile abalones (Liang et al., 2014).
The results of the present study showed the successful confirmation of hybrid status between H. discus hannai and H. gigantea using RAPD markers and the early survival and growth traits of these hybrids. Considering that hybridization is particularly useful if the heritability of a trait is low, the intrspecies selection may be inefficient (Elliot, 2000). Some underpinning strategic applied research should be conducted to investigate further the hybrid between H. discus hannai and H. gigantea abalones. From the results of this study, a breeding strategy using hybrids will be available for future abalone culture that is especially applicable to warm temperature water areas. The traits intermediate between parents would be useful for selective breeding material. Furthermore, given that H. discus hannai is the most popular and best priced abalone product in the international market, due to the quality of its meat and traditional consumer preferences (Cook and Gordon, 2010), the selection of specific hybrid genotypes could preserve the phenotypic attributes of the meat of H. discus hannai, such as flavor, tenderness and texture, as well as the biotechnological solutions to improve fertilization rates of hybrid abalones, given its potential to diversify the Korean abalone industry.
- References
-
1. Ahmed F, Koike Y, Strussmann CA, Yamasaki I. Yokota M, Watanabe S. 2008. Genetic characterization and gonad development of artificially produced interspecific hybrids of the abalones, Haliotis discus discus Reeve, Haliotis gigantean Gmelin and Haliotis madaka Habe. Aquacul Res 39: 532-541.


-
2. An HS, Jee YJ, Han SJ, Kim BL, Kim EM, Park IS. 2007. Induction of a New Hybrid between Haliotis gigantea Gmelin (♀) and H. discus discus Reeve (♂). Korean J Genetics 29: 239-244.
-
3. An HS, Jee YJ, Min KS, Kim BL, Han SJ. 2005. Phylogenetic analysis of six species of Pacific abalone (Haliotidae) based on DNA sequences of 16s rRNA and cytochrome c oxidase subunit Ⅰ mitochondrial genes. Mar Biotechnol 7: 373-380.
-
4. Asahida T, Kobayashi T, Saitoh K, Nakayama I. 1996. Tissue preservation and total DNA extraction from fish stored at ambient temperature using buffers containing high concentration of urea. Fisher Sci 62: 727-730.
-
5. Bartley DM, Rana K, Immink AJ. 2001. The use of inter-specific hybrids in aquaculture and fisheries. Rev Fish Biol Fish 10: 325-337.
-
6. Choi CG, Kim JM. 2012. Detection of Laminariaceae Species Based on PCR by Family-specific ITS Primers. Fish Aquat Sci 2: 157 -162.
-
-
8. Dang VT, Speck P, Benkendorff K. 2012. Influence of elevated temperatures on the immune response of abalone, Haliotis rubra. Fish Shellfish Immunol 32: 732-740.
-
9. Diaz F, del Rio-Portilla MA, Sierra E, Aguilar M, Re-Araujo AD. 2000. Preferred temperature and critical thermal maxima of red abalone Haliotis rufescens. J Thermal Bio 25: 257-261.
-
10. Elliott NG. 2000. Genetic improvement programmes in abalone: what is the future? Aquacult Res 31: 51-59.
-
11. Gao SS, Wang Q, Wang RB, Li X. 2000. Abalone. Liaoning Science and Technology Press, Shenyang, China, 14pp (in Chinese).
-
12. Geiger DL, Poppe GT. 2000. A Conchological Iconography. In The Family Haliotidae, Geiger DL and Poppe GT, eds., Conch Books, Hackenheim, Germany, pp 135.
-
-
14. Hulata G. 2001. Genetic manipulations in aquaculture: a review of stock improvement by classical and modern technologies. Genetica 111: 155-173.
-
15. Ibarra AM, Harnandez-Ibarra NK, Cruz P, Perez-Enriquez R, Avila S, Ramirez JL. 2005. Genetic certification of presumed hybrids of blue × red abalone (Haliotis fulgens Philippi and H. rufescens Swainson). Aquacult Res 36: 1-13.
-
16. Jana TK, Singh NK, Koundal KR, Sharma TR. 2005. Genetic differentiation of charcoal tot pathogen, Macrophomina phaseolina, into specific groups using URP-PCR. Can J Microbiol 51: 159 -164.
-
17. Kang HW, Go SJ, Eun MY, Chung TY. 1997. Genetic characterization of repetitive sequence derived from Korean red rice. Proceeding of the 8th SABRAO general congress and the Annual Meeting of the Korean, 24-28 September, Seoul, Korea, 421 -422.
-
18. Kang HW, Park DS, Go SJ, Eun MY. 2002. Fingerprinting of diverse genomes using PCR with universal rice primers generated from repetitive sequence of Korean weedy rice. Mol Cells 13: 281-287.
-
19. Kim Y, Jee YJ, Kim SH, Baik JM, Yang SG. 1988. Interspecific characteristic of abalones in the southern waters of Korea. Bull Nat Fish Res Dev Agency 42: 71-80.
-
20. Klinbunga S, Ampayup P, Leelatanawit R, Tassanakajon A, Hirono I, Auki T, Jarayabhand P, Menasveta P. 2004. Species identifi- cation of the tropical abalone (Haliotis asinine, Haliotis ovina and Haliotis varia) in Thailand using RAPD and SCAR markers. J Bioch Mol Biol 37: 213-222.
-
21. Klinbunga S, Ampayup P, Tassanakajon A, Jarayabhand P, Yoosukh W. 2000. Development of species-specific markers of the tropical oyster (Crassostrea belcheri) in Thailand. Mar Biotechnol 2: 476-484.
-
22. Korea National Statistical Office. 2012. KOSIS statistical DB. Daejeon, Korea. Available at http://kostat.go.kr.
-
23. Lafarga-de la Cruz F, Gallardo-Escarate C. 2011. Interspecies hybrids in Haliotis: natural and experimental evidence and its impact on abalone aquaculture. Rev Aquacult 3: 74-99.
-
24. Liang S, Luo X, You W, Luo L, Ke C. 2014. The role of hybridization in improving the immune response and thermal tolerance of abalone. Fish Shellfish Immunol 39: 69-77.
-
25. Liu ZJ, Cordes JF. 2004. DNA marker technologies and their applications in aquaculture genetics. Aquaculture 238: 1-37.
-
26. Marín SA, Haye PA, Marchant S, Winkler FM. 2007. Molecular markers used to analyze species-specific status in abalones with ambiguous morphology. J Shellfish Res 3: 833-837.
-
27. Rajora OP, Rahman MH. 2003. Microsatellite DNA and RAPD fingerprinting, identification and genetic relationships of hybrid poplar (Populus × Canadensis) cultivars. Theor Appl Genet 106: 470-477.
-
28. Williams JKG, Kubelik AR, Livak KJ, Rafalsky JA, Tyngey SV. 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res 18: 6531-6535.
-
Copyright (C) 2024. The Korean Society of Marine Life Science