










JMLS 2024 June;9(1):33-40. 10.23005/ksmls.2024.9.1.33 Epub 2024 June 13
Copyright © 2024 by The Korean Society of Marine Life Science
Exposure to Triclosan Induces Mortality through Oxidative Stress and DNA Damage in the Java Medaka Oryzias javanicus
Seong Duk Do; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea
Jae-Sung Rhee; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea; Research Institute of Basic Sciences, Incheon National University, Incheon 22012, Korea; Yellow Sea Research Institute, Incheon 22012, Korea
- Abstract
To understand the detrimental effects of triclosan on Java medaka (Oryzias javanicus) embryos, fertilized embryos were exposed to different concentrations (1, 10, 50, 100, 200, 400, 600, 800, and 1,000 μg l-1) of triclosan until hatching. Then, we examined the survival rate and developmental parameters as well as alterations in antioxidant constituents and DNA damage markers. The results showed dose-dependent mortality, hatching delays, and developmental abnormalities in the embryos. Additionally, there were significant increases in oxidative stress parameters and antioxidant responses, along with elevated DNA damage. These findings suggest that sublethal concentrations of triclosan induce toxic effects through oxidative stress on Java medaka embryos, as evidenced by changes in in vivo parameters and biochemical constituents.
Keywords: Triclosan Java medaka Embryo toxicity Oxidative stress DNA damage
Correspondence to: Jae-Sung Rhee; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea
- Received
- 17 April 2024;
- Revised
- 21 April 2024;
- Accepted
- 27 May 2024.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

Introduction
Triclosan, a chlorinated aromatic hydrocarbon containing anti- mycotic and antimicrobial functional groups such as ether and phenol, is widely used in various products including cosmetics (e.g., fragrances, shampoos, and moisturizers), drugs (e.g., isoniazid and diazoborine), and general health products (e.g., toothpaste, acne treatment, skin cleanser, and lotion) (Ahn et al., 2008; Dhillon et al., 2015). Its extensive usage, easy release into the environment, and incomplete removal through wastewater treatment contrib- ute to its frequent detection in different environmental matrices such as effluents, surface water, drinking water, and groundwater (Halden and Paull, 2005; Ramaswamy et al., 2011). Triclosan can bioaccumulate in aquatic food chains, particularly through fat tissues, moving from lower to higher trophic levels due to its relatively high lipophilicity, as indicated by a high log Kow of 4.8 (Orvos et al., 2002). The bioaccumulation of triclosan presents risks to non-target organisms within aquatic ecosystems. Its widespread presence in waterbodies and potential toxicity to aquatic organisms raise significant global environmental concerns.
The attention given to the toxicological risks of triclosan for various aquatic organisms has been substantial (Orvos et al., 2002; Kumar et al., 2021). As a result, bioconcentration, bioaccumulation, and evident toxicity of triclosan have been consistently reported across a variety of aquatic animals (Oliveira et al., 2009; Dann and Hontela, 2011). Triclosan basically operates by inhibiting fatty acid synthesis through the reduction of the enzyme enoyl-acyl carrier protein reductase (DeLorenzo et al., 2008). Acute toxicity due to triclosan exposure has been highlighted in numerous aquatic organisms (Orvos et al., 2002; Kumar et al., 2021; Do et al., 2024). Particularly in the genus Oryzias of fish, toxicity levels have been documented within a range from micrograms to milligrams, with these values closely associated with survival rates, impaired growth, and biochemical modulations (Ishibashi et al., 2004; Kim et al., 2009; Nassef et al., 2010; Horie et al., 2018; Mihaich et al., 2019; Song et al., 2020).
Exposure to xenobiotics, whether through direct contact or indirect pathways, can induce changes in the physiological and biochemical processes of aquatic organisms (Livingstone, 2001). These shifts in developmental parameters exhibited by such organisms can serve as sensitive indicators, providing early warn- ings of the harmful effects of contaminants in aquatic environ- ments. Previous studies have shown that exposure to triclosan can result in oxidative stress, disruption of hormone homeostasis, and apoptosis among the biochemical processes (Ruszkiewicz et al., 2017; Kumar et al., 2021). Oxidative stress is an event caused for the imbalance between the levels of reactive oxygen species (ROS; e.g., hydroxyl radicals, superoxide radicals, and hydrogen peroxide) and antioxidants (Lushchak, 2011). Typically, ROS attack cellular membrane lipids, resulting in the production of malondialdehyde (MDA) and prompt antioxidant responses within cells, owing to lipid peroxidation, DNA damage, and protein oxidation (Lesser, 2006). MDA serves as an indicator of lipid peroxidation, high- lighting oxidative damage that occurs when organisms are unable to counteract oxidative stress effectively through the utilization of antioxidant defenses. Antioxidant components, including catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx), and glutathione S-transferase (GST), play a vital role in neutralizing ROS and reducing the risk of oxidative damage (Sies, 1991).
Since there is limited data on the harmful effects of triclosan on the embryonic development of marine fish, although its con- sistent detection in marine ecosystems, we have hypothesized its detrimental effects on embryonic development in this study. The Java medaka (Oryzias javanicus) was utilized as a non-target fish model to investigate the potentially harmful effects of triclosan. Java medaka are commonly found in coastal areas, where environ- mental pollution is prevalent. These fish are particularly vulnerable to various exogenous xenobiotics during their embryonic and larval stages (Nam et al., 2020). First, we examined how triclosan affects the survival and developmental progress of Java medaka embryos from the start to the end of the embryonic stage, in- cluding morphometric analysis on their fin, spinal, and cardiac development. We then theorized that oxidative stress plays a significant role in influencing changes in embryo damage levels. Consequently, we analyzed biochemical parameters related to oxidative status, along with DNA stability parameters, in the ex- posed embryos. The results will contribute to determining whether triclosan has harmful effects on the embryonic development of non-target marine fish.
Materials and Methods
1. Java medaka
The Java medaka Oryzias javanicus were maintained under controlled conditions at 26℃ with a light-dark cycle of 14 hours of light and 10 hours of darkness in artificial seawater (TetraMarine Salt Pro, TetraTM, Cincinnati, OH, USA), with an oxygen concen- tration of 6.53 ± 0.67 mg O2 l-1, adjusted to 12 practical salinity units. The fish were housed in glass aquaria with a capacity of 60 liters each, equipped with continuous aeration. Each aquarium housed up to 30 adult fish of both sexes. They were fed Artemia salina (< 6 hours after hatching) twice daily until satiation. All animal handling and experimental procedures followed the guide- lines of the Animal Welfare Ethical Committee and the Animal Experimental Ethics Committee of Incheon National University (Incheon, South Korea).
2. Exposure
Triclosan was obtained from Sigma-Aldrich Co. (St. Louis, MO, USA). A stock solution of triclosan (100 mg ml-1) was prepared by dissolving it in dimethyl sulfoxide (DMSO) from Sigma-Aldrich Co. The maximum concentration of DMSO was 0.02%. An equal volume of DMSO was added to the solvent control group. The stock solution was stored in dark conditions until treatment to prevent photodissociation. Working solutions were prepared by diluting each stock solution in 0.22 μm-filtered medium.
Naturally fertilized embryos, 3 hours post-fertilization confirmed with microscopy analysis, were exposed to nine concentrations (1, 10, 50, 100, 200, 400, 600, 800, and 1,000 μg l-1) until hatching. The experimental conditions were consistent, with the control group exposed to DMSO. In a 100 ml aquarium volume, a total of 90 embryos were exposed to each concentration. After expo- sure, the specimens were divided into three replicates, with each replicate containing 30 embryos per concentration (n = 30). The triclosan solutions were renewed daily until hatching, with deceased embryos promptly removed from the aquarium. Survival rate and hatching duration were evaluated by monitoring developmental traits throughout embryonic progression. For the biochemical assay, the fish were anesthetized with MS-222 solution (200 mg/l tricaine methanesulfonate, Sigma-Aldrich, St. Louis, MO, USA).
3. Measurement of biochemical parameters
Biochemical parameters were assessed using viable embryos from samples exposed to triclosan concentrations up to 100 μg l-1. Beyond this threshold, significant mortality occurred, increasing the risk of protein damage and elevated error rates in the samples.
Intracellular levels of ROS and MDA contents were evaluated following established methodologies outlined in our prior inves- tigations involving Java medaka embryos (Nam et al., 2020). After exposure to varying concentrations of triclosan, fully developed embryos (approximately 168 hours post-fertilization) were col- lected (n = 50 for each concentration) and homogenized. The homogenates were then centrifuged at 10,000 × g for 20 minutes at 4℃, and the resulting supernatants were retrieved for analysis. In 96-well black plates filled with phosphate-buffered saline buffer, the supernatant, along with the probe (H2DCF-DA at a final con- centration of 40 μM), was added to achieve a final volume of 200 μl. Measurements were carried out at an excitation wave- length of 485 nm and an emission wavelength of 520 nm using a ThermoTM Varioskan Flash spectrophotometer (Thermo Fisher Scientific, Tewksbury, MA, USA), with normalization performed based on total protein values determined using the Bradford method.
The procedure for measuring MDA contents involved several steps. Initially, the pooled embryos were homogenized using a Teflon homogenizer in Tris buffer (20 mM). Subsequently, the homogenates underwent centrifugation at 30,000 × g at 4℃ for 30 minutes, and the resulting supernatants were subjected to heat denaturation at 75℃ for 15 minutes. Thiobarbituric acid reactive substances were quantified at 535 nm using a Thermo Varioskan Flash spectrophotometer (Thermo Fisher Scientific). The concentrations of these substances were determined using a standard curve established with MDA bis(dimethylacetal) (Sigma-Aldrich, Inc., St. Louis, MO, USA). Finally, MDA levels were calcu- lated based on a calibration curve and expressed as nM of MDA per μg of the total sample.
The antioxidant defense system's enzyme activities, such as GST, GPx, CAT, and SOD, were evaluated following methodologies described in our prior study on Java medaka embryos (Nam et al., 2020). For GST activity, the samples were homogenized using a Teflon homogenizer with cold buffer [1:4, w/v; containing 0.25 M sucrose, 10 mM Tris, 1 mM ethylenediaminetetraacetic acid (EDTA), 0.2 mM dithiothreitol (DTT), and 0.1 mM phenylmethylsulfonyl fluoride (PMSF), pH 7.4], and then centrifuged at 10,000 g for 10 minutes at 4°C. The resulting supernatant, which contained the enzyme, was collected for enzymatic analysis using 1-chloro-2,4-dinitrobenzene (CDNB) as a substrate. GST activity was measured at 340 nm at 25℃. Samples were washed and homogenized using a Teflon homogenizer, then centrifuged at 3,000 to 10,000 × g for 10 minutes at 4℃. The resultant upper aqueous layer was used for enzymatic assays, following the manufacturer's instructions. GPx activity was determined using a glutathione peroxidase cellular assay kit (Sigma-Aldrich, Inc.), while CAT and SOD activities were measured using catalase (Sigma-Aldrich Chemie, Switzerland) and SOD assay kits (Sigma-Aldrich Chemie), respectively, at 25℃ with absorbance readings at 520 nm and 440 nm. Oxidative and anti- oxidant parameters were quantified using a ThermoTM Varioskan Flash spectrophotometer, with normalization based on total protein values and expressed as a percentage of the control. Total protein concentrations were determined using the Bradford method.
4. DNA stability
DNA damage assessment was conducted using the alkaline single-cell gel electrophoresis (Comet) assay, following method- ologies established in our previous studies (Shin et al., 2019; Nam et al., 2020). Samples were diluted with cold fetal bovine serum (Gibco, MA, USA) at 4℃ and mixed with 0.5% low melting-point (LMP) agarose. The mixture was applied onto slides pre-coated with 1% normal melting-point agarose and covered with cover- slips. After solidification for 10 minutes at 4℃, 0.5% LMP agarose was added to cover the slides, followed by immersion in cold lysing solution (2.5 M NaCl, 100 mM Na2-EDTA, 10 mM Tris, pH 10, 10% DMSO, and 1% Triton X-100) at 4℃ for 2 hours. Sub- sequently, the slides were placed in unwinding buffer (1 mM Na2-EDTA, 300 mM NaOH, pH 13.5) for 20 minutes, followed by electrophoresis at 25 V and 300 mA for 25 minutes. Post-electrophoresis, the slides were washed three times in 0.4 M Tris buffer for 10 minutes each, dehydrated in 70% ethanol, and stained with ethidium bromide (20 μg ml-1). A total of 100 cells were randomly observed under a fluorescent microscope (Nikon, Tokyo, Japan), and images were analyzed using Komet 6.0 soft- ware (Kinetic Imaging, Liverpool, UK). The olive tail moment (tail DNA% multiplied by tail moment length), as determined by the software, was utilized to quantify DNA stability.
5. Statistical analysis
Statistical analysis was performed using the SPSS statistical software program (version 17.0; SPSS Inc., Chicago, IL, USA), and data were expressed as mean ± standard deviation (S.D). To deter- mine significant differences between control and treatment groups, one-way analysis of variance (ANOVA) followed by Tukey's test was employed at a significance level of p < 0.05.
Results and Discussion
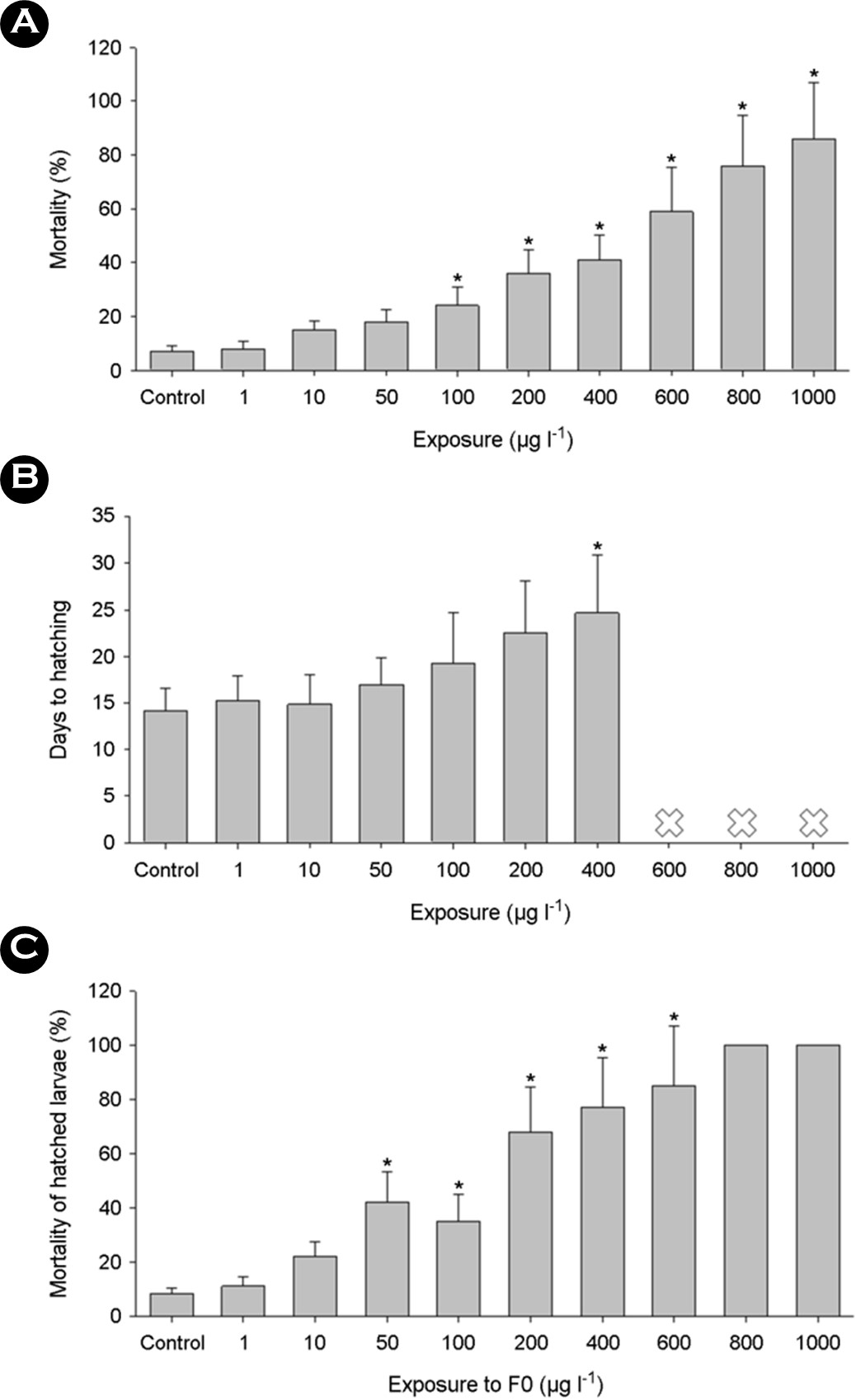
The mortality of embryos increased in a dose-dependent manner, along with notable delays in hatching when exposed to triclosan (p < 0.05) (Figs. 1A and 1B). Examination of survival rates upon exposure to triclosan revealed a critical threshold for embryo survival. For instance, the 96-hour LC50 values were recorded at 399 μg l-1 for embryos and 600 μg l-1 for larvae in Japanese medaka (Ishibashi et al., 2004; Kim et al., 2009). In zebrafish embryo, 96 h-LC50 value was 420 μg l-1 (Oliveira et al., 2009). Larvae of Java medaka, hatched from surviving embryos, exhibited adverse effects from prior exposure to triclosan during embryonic stages (p < 0.05) (Fig. 1C), despite being raised in clean artificial seawater without triclosan treatment. Overall mortality rates surpassed those of F0 mortality. Shortly after hatching, a sharp rise in mortality was noted, alongside dose-dependent mortality in Japanese medaka embryos exposed to triclosan (Horie et al., 2018). The observed mortality and delayed hatching in embryos suggest vulnerability of earlier developmental stages to external triclosan exposure. Furthermore, these findings suggest that even sub-lethal concen- trations of triclosan exposure could lead to significant mortality in fish larvae post-hatching.
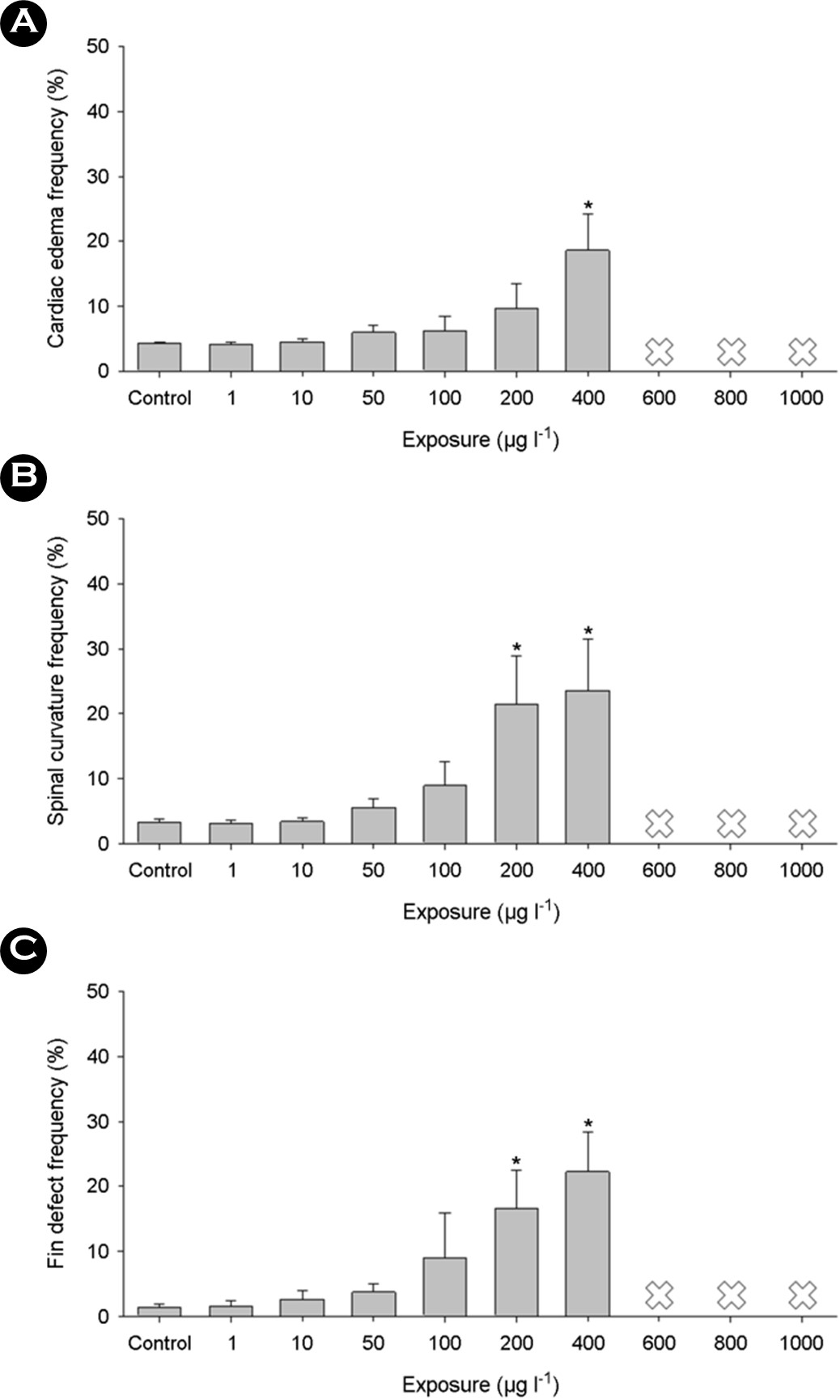
Increasing concentrations of triclosan during early develop- mental stages led to significant malformations, including cardiac edema (Fig. 2A), spinal curvature (Fig. 2B), and fin defects (Fig. 2C) (p < 0.05). These findings suggest that triclosan exposure induced abnormal developments, contributing to embryo mortality. Although fish-related data are limited, a previous study demon- strated notable adverse effects of triclosan on the embryonic development of Japanese medaka. Developmental exposure to triclosan resulted in an enlarged yolk sac, decreased head trunk angle, and severe edema in the pericardial region (Song et al., 2020). Severe edema with reduced heartbeat induced by develop- mental exposure to triclosan has also been observed in zebrafish (Saley et al., 2016; Macedo et al., 2017). Triclosan-exposed zebrafish exhibited delayed hatching, along with delays in otolith formation, and pigmentation of the eyes and body, accompanied by pine malformations and pericardial edema (Oliveira et al., 2009).
Exposure to triclosan resulted in a notable increase in intra- cellular ROS and MDA levels (p < 0.05) (Figs. 3A and 3B). MDA is typically produced as a final byproduct of lipid peroxidation and/or mitochondrial dysfunction, processes intricately associated with ROS generation (Lushchak, 2011). Hence, MDA serves as a biomarker of lipid peroxidation, indicating oxidative damage when organisms fail to adequately counter oxidative stress through antioxidant mechanisms (Lesser, 2006). The susceptibility of embry- onic stages in aquatic animals to oxidative stress has been well-documented (Winston and Di Giulio, 1991; Valavanidis et al., 2006). Triclosan significantly elevated MDA content in the liver tissue of adult zebrafish (Gyimah et al., 2020). Additionally, acute exposure to triclosan markedly increased ROS levels in zebrafish embryos (Parenti et al., 2019; Liu et al., 2022). Considering these findings, the compromised responses of ROS and MDA observed in Java medaka are likely associated with the heightened mortality observed in embryos exposed to triclosan.
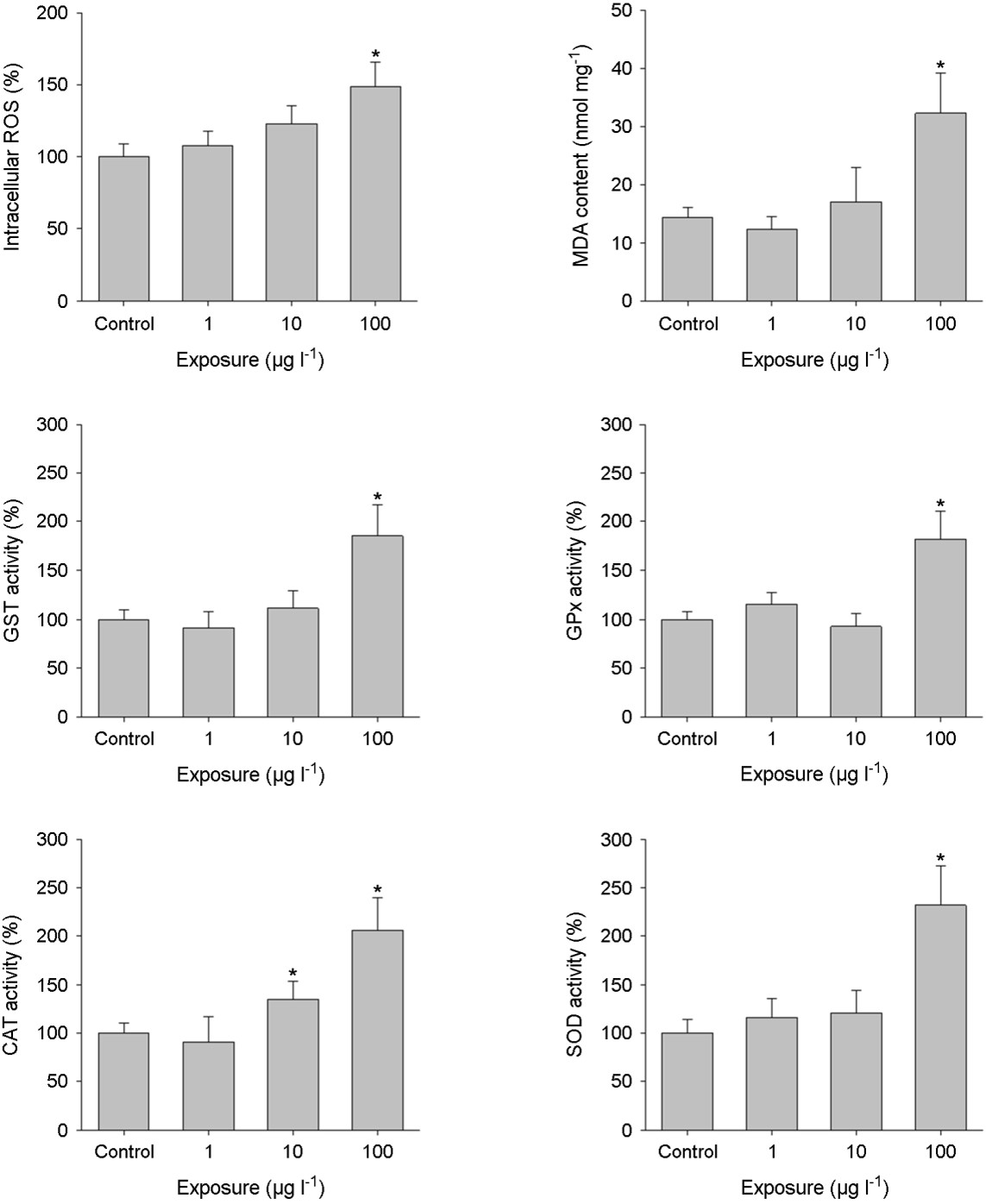
Aquatic organisms have evolved antioxidant defense systems to combat external stressors and alleviate damage induced by ROS (Winston and Di Giulio, 1991; Valavanidis et al., 2006; Nam et al., 2023). Excessive oxidative stress can disrupt this defense system by altering the levels or activity of antioxidant components (Lushchak, 2011). Exposure to triclosan significantly increased the levels of antioxidant enzymatic activities in Java medaka (p < 0.05) (Figs. 3C, 3D, 3E, and 3F), indicating an amplified total oxyradical scavenging capacity in the embryos, triggered by the oxidative stress provoked by triclosan exposure. GST plays a pivotal role as a metabolic enzyme, working alongside GSH to facilitate the detoxification of both internally generated and externally encoun- tered toxic substances, thereby alleviating damage caused by oxidative stress (Valavanidis et al., 2006). GPx has a critical func- tion in neutralizing hydrogen and lipid peroxides in the presence of GSH, thus bolstering the oxidative defense system by safe- guarding cells against apoptosis and scavenging oxidative radicals (Lushchak, 2011). CAT and SOD are fundamental constituents of cellular defense mechanisms against oxidative stress (Lesser, 2006). SOD facilitates the dismutation of superoxide anions into hydrogen peroxide and molecular oxygen, while CAT subsequently converts hydrogen peroxide into water and oxygen (Valavanidis et al., 2006). Therefore, the activation of the antioxidant defense system indi- cates the embryos' capacity to maintain balance in response to oxidative stress induced by triclosan exposure. Similarly, a parallel increase in the antioxidant defense system response has been observed in zebrafish following exposure to triclosan. Exposure to sub-lethal concentrations of triclosan markedly elevated the enzy- matic activities of GST, GPx, CAT, and SOD in zebrafish embryos (Parenti et al., 2019).
The exploration of triclosan's potential impact on DNA stability offered corroborative evidence for the hypothesis proposing that triclosan triggered the generation of intracellular ROS. This was evidenced by the significantly heightened levels of DNA frag- mentation compared to control levels (p < 0.05) (Fig. 4A). Simi- larly, there were noteworthy rises in the percentage of tail DNA subsequent to exposure to triclosan (p < 0.05) (Fig. 4B). Dose-dependent DNA damage caused by triclosan was reported in the hepatocytes of adult zebrafish (Gyimah et al., 2020). Triclosan-exposed zebrafish embryos exhibited a significant increase in cell necrosis or apoptosis due to substantial DNA damage (Parenti et al., 2019; Liu et al., 2022). The increased levels of DNA damage observed following triclosan exposure aligned with increased mortality rates and oxidative stress.

In summary, exposure to triclosan via waterborne pathways led to a significant uptick in both mortality and the occurrence of developmental abnormalities. This result can be linked to induced oxidative stress, despite the protective presence of the chorion and membrane. Although embryos demonstrated resilience to sublethal levels of triclosan, the inability to uphold molecular and biochemical balance could result in significant mortality and abnormal embryonic development in fish. Our findings illuminate the adverse effects of triclosan and underscore the necessity of stricter regulations to minimize its release into aquatic environ- ments, thereby enabling more precise risk assessment procedures.
- References
-
1. Ahn KC, Zhao B, Chen J, Cherednichenko G, Sanmarti E, Denison MS, Lasley B, Pessah IN, Kültz D, Chang DPY, Gee SJ, Hammock BD. 2008. In vitro biologic activities of the anti- microbials triclocarban, its analogs, and triclosan in bioassay screens: receptor-based bioassay screens. Environ Health Perspect 116: 1203-1210.
-
2. Dann AB, Hontela A. 2011. Triclosan: environmental exposure, toxicity and mechanisms of action. J Appl Toxicol 31: 285-311.
-
3. DeLorenzo ME, Keller JM, Arthur CD, Finnegan MC, Harper HE, Winder VL, Zdankiewicz DL. 2008. Toxicity of the antimicrobial compound triclosan and formation of the metabolite methyl-triclosan in estuarine systems. Environ Toxicol 23: 224-232.
-
4. Dhillon GS, Kaur S, Pulicharla R, Brar SK, Cledón M, Verma M, Surampalli RY. 2015. Triclosan: current status, occurrence, environmental risks and bioaccumulation potential. Int J Environ Res Public Health 12: 5657-5684.
-
5. Do SD, Haque MN, Kim J, Im D-H, Rhee J-S. 2024. Acute and chronic effects of triclosan on the behavior, physiology, and multigenerational characteristics of the water flea Moina macrocopa. Comp Biochem Physiol C 276: 109810.
-
6. Gyimah E, Dong X, Qiu W, Zhang Z, Xu H. 2020. Sublethal con- centrations of triclosan elicited oxidative stress, DNA damage, and histological alterations in the liver and brain of adult zebrafish. Environ Sci Pollut Res 27: 17329-17338.
-
7. Halden RU, Paull DH. 2005. Co-occurrence of triclocarban and triclosan in US water resources. Environ Sci Technol 39: 1420-1426.
-
8. Horie Y, Yamagishi T, Takahashi H, Iguchi T, Tatarazako N. 2018. Effects of triclosan on Japanese medaka (Oryzias latipes) during embryo development, early life stage and reproduc- tion. J Appl Toxicol 38: 544-551.
-
9. Ishibashi H, Matsumura N, Hirano M, Matsuoka M, Shiratsuchi H, Ishibashi Y, Takao Y, Arizono K. 2004. Effects of triclosan on the early life stages and reproduction of medaka Oryzias latipes and induction of hepatic vitellogenin. Aquat Toxicol 67: 167-179.
-
10. Kim J-W, Ishibashi H, Yamauchi R, Ichikawa N, Takao Y, Hirano M, Koga M, Arizono K. 2009. Acute toxicity of pharmaceutical and personal care products on freshwater crustacean (Tham- nocephalus platyurus) and fish (Oryzias latipes). J Toxicol Sci 34: 227-232.
-
11. Kumar S, Paul T, Shukla SP, Kumar K, Karmakar S, Bera KK. 2021. Biomarkers-based assessment of triclosan toxicity in aquatic environment: A mechanistic review. Environ Pollut 286: 117569.
-
12. Lesser MP. 2006. Oxidative stress in marine environments bio- chemistry and physiological ecology. Annu Rev Physiol 68: 253-278.
-
13. Liu F, Zhang Y, Wang F. 2022. Environmental relevant concen- trations of triclosan affected developmental toxicity, oxidative stress, and apoptosis in zebrafish embryos. Environ Toxicol 37: 848-857.
-
14. Livingstone D. 2001. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organ- isms. Mar Pollut Bull 42: 656-666.
-
15. Lushchak VI. 2011. Environmentally induced oxidative stress in aquatic animals. Aquat Toxicol 101: 13-30.
-
16. Macedo S, Torres T, Santos MM. 2017. Methyl-triclosan and triclosan impact embryonic development of Danio rerio and Paracentrotus lividus. Ecotoxicology 26: 482-489.
-
17. Mihaich E, Capdevielle M, Urbach-Ross D, Gallagher S, Wolf J. 2019. Medaka (Oryzias latipes) multigeneration test with triclosan. Environ Toxicol Chem 38: 1770-1783.`
-
18. Nam S-E, Saravanan M, Rhee J-S. 2020. Benzo[a]pyrene con- strains embryo development via oxidative stress induction and modulates the transcriptional responses of molecular biomarkers in the marine medaka Oryzias javanicus. J Environ Sci Health A Tox Hazard Subst Environ Eng 55: 1050-1058.
-
19. Nam S-E, Bae D-Y, Ki J-S, Ahn C-Y, Rhee J-S. 2023. The importance of multiomics approaches for the health assessment of freshwater ecosystems. Mol Cell Toxicol 19: 3-11.
-
20. Nassef M, Matsumoto S, Seki M, Khalil F, Kang IJ, Shimasaki Y, Oshima Y, Honjo T. 2010. Acute effects of triclosan, diclofenac and carbamazepine on feeding performance of Japanese medaka fish (Oryzias latipes). Chemosphere 80: 1095-1100.
-
21. Oliveira R, Domingues I, Grisolia CK, Soares AM. 2009. Effects of triclosan on zebrafish early-life stages and adults. Environ Sci Pollut Res 16: 679-688.
-
22. Parenti CC, Ghilardi A, Della Torre C, Mandelli M, Magni S, Del Giacco L, Binelli A. 2019. Environmental concentrations of triclosan activate cellular defence mechanism and generate cytotoxicity on zebrafish (Danio rerio) embryos. Sci Total Environ 650: 1752-1758.
-
23. Ramaswamy BR, Shanmugam G, Velu G, Rengarajan B, Larsson DGJ. 2011. GC-MS analysis and ecotoxicological risk assess- ment of triclosan, carbamazepine and parabens in Indian rivers. J Hazard Mater 186: 1586-1593.
-
24. Ruszkiewicz JA, Li S, Rodriguez MB, Aschner M. 2017. Is Triclosan a neurotoxic agent? J Toxicol Environ Health B Crit Rev 20: 104-117.
-
25. Saley A, Hess M, Miller K, Howard D, King-Heiden TC. 2016. Car- diac toxicity of triclosan in developing zebrafish. Zebrafish 13: 399-404.
-
26. Shin YK, Nam S-E, Kim WJ, Seo DY, Kim Y-J, Rhee J-S. 2019. Red tide dinoflagellate Cochlodinium polykrikoides induces sig- nificant oxidative stress and DNA damage in the gill tissue of the red seabream Pagrus major. Harmful Algae 86: 37-45.
-
27. Sies H. 1991. Oxidative stress: from basic research to clinical application. Am J Med 91: S31-S38.
-
28. Song X, Wang X, Bhandari RK. 2020. Developmental abnor- malities and epigenetic alterations in medaka (Oryzias latipes) embryos induced by triclosan exposure. Chemosphere 261: 127613.
-
29. Valavanidis A, Vlahogianni T, Dassenakis M, Scuollos M. 2006. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol Environ Saf 64: 178-189.
-
30. Winston GW, Di Giulio RT. 1991. Prooxidant and antioxidant mech- anisms in aquatic organisms. Aquat Toxicol 19: 137-161.
-
31. Orvos DR, Versteeg DJ, Inauen J, Capdevielle M, Rothenstein A, Cunningham V. 2002. Aquatic toxicity of triclosan. Environ Toxicol Chem 21: 1338-1349.
Copyright (C) 2025. The Korean Society of Marine Life Science