










JMLS 2023 December;8(2):186-189. 10.23005/ksmls.2023.8.2.186 Epub 2023 December 17
Copyright © 2023 by The Korean Society of Marine Life Science
Analysis of UCP1 Expression in Rainbow Trout Gonadal Cell Line RTG-2 Indicates its Marginal Response to Adipogenic Inducers Compared to Mammalian Cell Lines
Sang-Eun Nam1, ; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea
Young-Joo Yun2, ; Research Institute of Basic Sciences, Incheon National University, Incheon 22012, Korea
Jae-Sung Rhee1,2, ; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea; Research Institute of Basic Sciences, Incheon National University, Incheon 22012, Korea
Hyoung Sook Park3*; Department of Song-Do Biological Engineering, Incheon Jaeneung University, Incheon 21987, Korea
- Abstract
Uncoupling protein 1 (UCP1) is a unique mitochondrial membranous protein expressed in brown adipose tissue (BAT) in mammals. While its expression in response to cold temperatures and adipogenic inducers is well-characterized in mammals and human infants, the molecular characterization and expression of UCP1 in fish remain unexplored. To address this gap, we analyzed UCP1 expression in response to adipogenic inducers in a fish cell line, rainbow trout gonadal cells (RTG-2), and compared it with UCP1 expression in three mammalian preadipocytes, 3T3-L1, T37i, and WT1 exposed to the Peroxisome proliferator-activated receptor gamma (PPARγ) agonists, rosiglitazone (Rosi). In mammalian preadipocytes, UCP1 protein was highly expressed by Rosi, with an induction of adipogenesis observed in a time-dependent manner. This suggests that UCP1 plays a significant role in adipogenesis in mammals. However, RTG-2 cells showed no response to adipogenic inducers and exhibited only marginal expressions of UCP1. These results imply that RTG-2 cells may lack crucial responsive mechanisms to adipogenic signals or that the adipogenic response is regulated by other mechanisms. Further studies are needed to confirm these phenomena in fish preadipocytes when an appropriate cell line is established in future research.
Keywords: Fish Adipogenesis UCP1 expression Rosiglitazone
Correspondence to: Hyoung Sook Park; Department of Song-Do Biological Engineering, Incheon Jaeneung University, Incheon 21987, Korea
- Received
- 15 October 2023;
- Revised
- 22 October 2023;
- Accepted
- 16 November 2023.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

In mammals, adipose tissues are typically classified into two main types: white adipose tissue (WAT) and BAT (Gesta et al., 2007). WAT, in general, serves as a repository for triglycerides, playing a crucial role in energy storage, maintaining glucose homeostasis, regulating inflammatory processes, and participating in endocrine functions (Billon and Dani, 2011). Conversely, the physiological role of BAT has been the subject of extensive investigation. Brown adipocytes within BAT are characterized by their heightened meta- bolic activity, which is facilitated by a large number of mito- chondria and numerous small triglyceride droplets (Cannon and Nedergaard, 2004). They are also known to possess specialized functions primarily centered around the expenditure of energy through non-shivering thermogenesis (NST) (Foster and Frydman, 1978; Himms-Hagen, 1984). This process generates heat energy either through the UCP1-dependent uncoupling of mitochondrial
respiration or via UCP1-independent signaling cascades (Cannon and Nedergaard, 2004; Wang et al., 2016). Furthermore, several studies have highlighted BAT's involvement in the endocrine system, as well as its role in maintaining glucose and lipid homeo- stasis (Stanford et al., 2012; Villarroya et al., 2017).
UCP1, a member of the mitochondrial carrier protein superfamily, is predominantly expressed in the inner mitochondrial mem- brane of BAT. Its primary function is to uncouple mitochondrial fatty acid oxidation from ATP production, achieving this by mediating a controlled release of the proton gradient generated by the respiratory chain (Ledesma et al., 2002; Ikeda and Yamada, 2020). This energy-dissipating mechanism is essential for NST and cold survival in various mammals, including rodents, hibernators, and human infants (Cannon and Nedergaard, 2004; Tine et al., 2012). The gene expression of UCP1 in BAT is mainly regulated by the upregulation or stimulation of several crucial components, such as PPARγ, β3Ad, and PGC1 (Seale et al., 2011; Boström et al., 2012; Park et al., 2016). In contrast to mammals, teleost fish has only WAT, not beige adipose tissue (BeAT) or BAT (Salmerón, 2018). However, UCP1 genes have been identified not only in mammals but also in fish (Jarmuszkiewicz et al., 1999; Jastroch et al., 2005), and it was reported that cold-water fish, such as rainbow trout lived below 21℃, contain a higher accumulation of unsaturated fatty acids for cold adaptation (Johnston, 1990; Crespi and New, 2009). To date, it remains unclear whether the classical mechanism of NST involving UCP1 exists in fish adapted to long-term low temperatures.
In this study, our objective was to investigate the potential for brown and/or beige adipocyte differentiation induced by PPARγ agonists in rainbow trout. Although there is limited research on adipogenic differentiation within BAT in fish, a recent report sug- gested the presence of adipocyte progenitors (APs) capable of adipogenic differentiation within gonadal WAT (Vishvanath and Gupta, 2019). Hence, we chose to perform our experiment with a gonadal cell line, RTG-2, derived from rainbow trout. Three types of preadipocytes (3T3-L1, T37i, and WT1), previously studies on brown or beige adipogenesis, and RTG-2 were treated with the PPARγ agonists. This approach allowed us to compare the results between the fish cell line RTG-2 and the mammalian cell lines.
The mouse preadipocyte cell lines, 3T3-L1 (CL-173), and RTG-2 (CCL-55), were procured from ATCC (Manassas, VA, USA). Mouse brown preadipocyte (SCC255) and mouse brown adipocyte (SCC- 250) were obtained from Sigma-Aldrich (St. Louis, MO, USA). To induce adipogenesis in the mouse preadipocyte cell lines, they were exposed to DMIT, as a mixture with 1 μM dexamethasone, 500 μM 3-Isobutyl-1-methylxanthine, 0.5 μg/mL insulin and 1 nM 3,3',5-Tniodo-L-thyronine, and/or 5 μM Rosi dissolved in culture media containing 10% FBS, for eight days. The culture media were refreshed every two days. A similar exposure regime was employed for the RTG-2 cells. Specifically, the RTG-2 cells were exposed to EMEM with DMIT and 5 μM Rosi. Additionally, cells were harvested, and total protein was extracted for the analysis of time-course UCP1 expression using immunoblotting. Cell samples were collected at day 0, 2, 4, and 8, and immediately fixed with 10% formalin for 20 min at room temperature for subsequent immunocytochemistry analysis and Oil Red O (ORO; O0625; Sigma-Aldrich) staining. Antibodies against UCP1 and β-tubulin were sourced from Abcam (ab10983; Cambridge, UK) and Cell Signaling Technology (2146S; Beverly, MA, USA), respectively. To visualize mitochondria, 1 μM MitoTracker® Deep Red FM (8778P; Cell Signaling Technology, Beverly, MA, USA) was applied to the cells for 30 min before fixation. For immunocytochemistry analysis, cells were subjected to permeabilization and background epitope blocking by incubation for 15 min in a blocking solution con- taining 2.5% BSA, 2% Normal Goat Serum, and 0.2% Triton X-100 in PBS. Subsequently, UCP1 as primary antibodies, diluted to 1:500 in blocking solution, were applied to the cells overnight at 4℃. After a 15-minute wash, secondary antibodies were intro- duced and allowed to incubate for 60 min at room temperature. Nuclei were visualized using DAPI (D9542; Sigma-Aldrich), and coverslips were mounted using Fluorescence Mounting Medium (S3023; Dako, Glostrup, Denmark). For ORO staining, the fixed cells were incubated for 1 h with 0.3% ORO in 60% isopropanol. Cell lysates, in a 2 × SDS sample buffer, were separated on SDS-polyacrylamide gels and transferred to Immobilon-P membranes (Millipore, Billerica, MA, USA). To block nonspecific proteins, mem- branes were treated with 5% skim milk in Tris-buffered saline containing 0.1% Tween 20 for 30 min. Subsequently, membranes were incubated overnight with primary antibodies diluted to 1:1,000 in the blocking solution. After this, the membranes were exposed to a horseradish peroxidase-conjugated secondary anti- body for 1 h in the blocking solution. Blots were visualized using the Clarity Western ECL Substrate kit (Bio-Rad, Hercules, CA, USA).
Immunocytochemistry analysis revealed that the three preadi- pocyte cell lines exhibited an increase in UCP1 expression, which coincided with the results obtained from mitochondrial membrane measurements (Fig. 1A-C). These increases were further validated by ORO staining results. Immunoblotting demonstrated a time-dependent increase in UCP1 expression in all three cell lines (Fig. 1D-F). However, in the case of RTG-2 cells, no significant differ- ences were observed in the potential induction of adipogenesis, as well as UCP1 expression (Fig. 2A). Additionally, there were no discernible morphological differences in RTG-2 cells following exposure to PPARγ agonists. ORO staining did not yield any positive results in RTG-2 cells. When we assessed the expression of UCP1 via immunoblotting, only marginal expressions were observed on each sampling day (Fig. 2B).
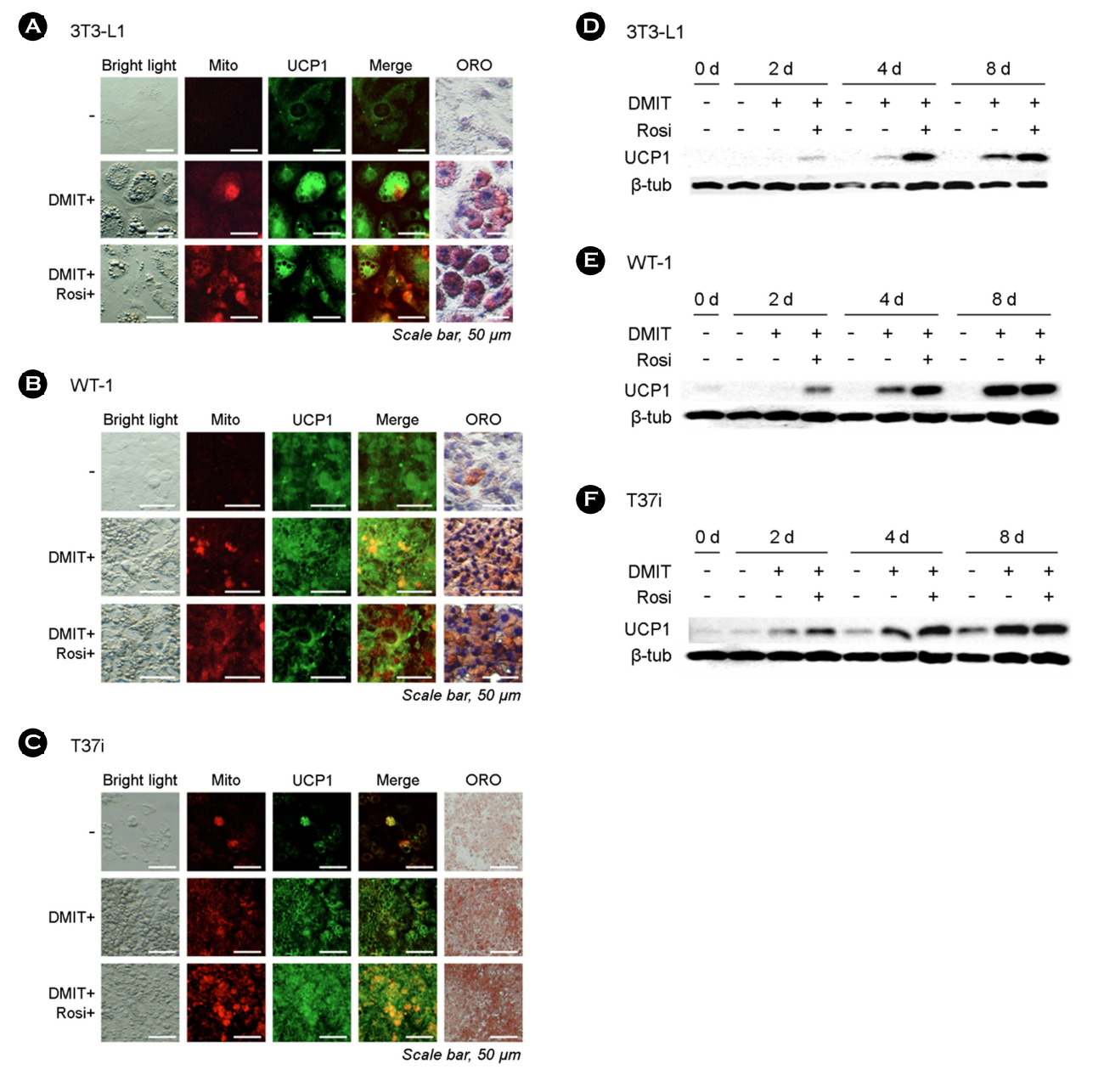
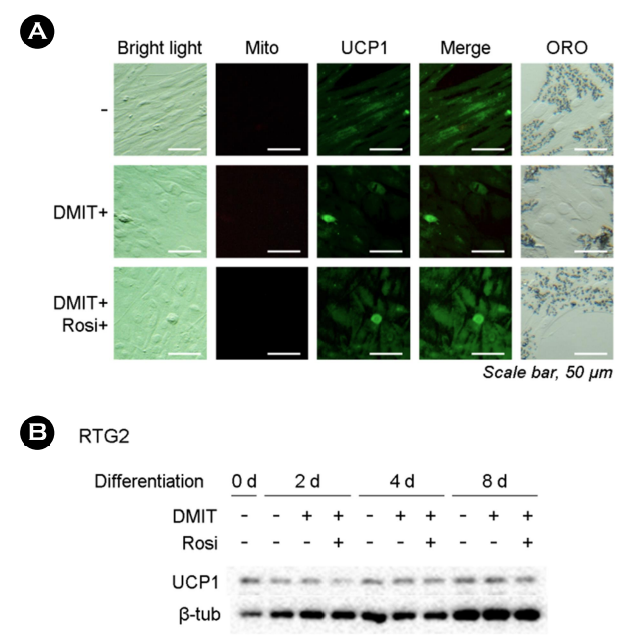
In summary, these findings suggest that the fish cell line RTG-2 may lack crucial responsive mechanisms to adipogenic signals, or that the adipogenic response is regulated by alternative mech- anisms. Given the high expression of UCP1 protein in mammalian preadipocytes induced by DMIT and/or Rosi, we can assume that our experimental conditions were appropriate to support the role of UCP1 in adipogenesis. However, a limitation of this study is the absence of suitable preadipocyte cell lines in fish and rainbow trout-specific or fish-specific UCP1 antibodies. To investigate the precise expression and function of UCP1, adipose stem cells, rather than gonadal cells, must be extracted from rainbow trout or other fish and differentiated into fat cells. Further research is necessary to confirm these phenomena in fish preadipocytes once an appropriate cell line becomes available in future studies.
- References
-
1. Billon N, Dani C. 2011. Developmental origins of the adipocyte lineage: new insights from genetics and genomics studies. Stem Cell Rev 8: 55-66.
-
2. Boström P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, Rasbach KA, Boström EA, Choi JH, Long JZ, Kajimura S, Zingaretti MC, Vind BF, Tu H, Cinti S, Højlund K, Gygi SP, Spiegelman BM. 2012. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 481: 463-468.
-
3. Cannon B, Nedergaard JAN. 2004. Brown adipose tissue: function and physiological significance. Physiol Rev 84: 277-359.
-
-
5. Foster DO, Frydman ML. 1978. Brown adipose tissue: the dominant site of nonshivering thermogenesis in the rat. In Effectors of Thermogenesis: Proceedings of a Symposium, Geneva, Switzerland, 14-16 Jul, 1977, pp 147-151 (in Switzerland).
-
6. Gesta S, Tseng YH, Kahn CR. 2007. Developmental origin of fat: tracking obesity to its source. Cell 131: 242-225
-
-
8. Ikeda K, Yamada T. 2020. UCP1 dependent and independent thermogenesis in brown and beige adipocytes. Front Endo- crinol 11: 498.
-
9. Jarmuszkiewicz W, Sluse-Goffart CM, Hryniewiecka L, Sluse FE. 1999. Identification and characterization of a protozoan un- coupling protein in Acanthamoeba castellanii. J Biol Chem 274: 23198-23202.
-
10. Jastroch M, Wuertz S, Kloas W, Klingenspor M. 2005. Uncoupling protein 1 in fish uncovers an ancient evolutionary history of mammalian nonshivering thermogenesis. Physiolgical Genomics 22: 150-156.
-
11. Johnston IA. 1990. Cold adaptation in marine organisms. Philos Trans R Soc B Biol Sci 326: 655-667.
-
12. Ledesma A, de Lacoba MG, Rial E. 2002. The mitochondrial un- coupling proteins. Genome Biology 3: 1-9.
-
13. Park HS, Ju UI, Park JW, Song JY, Shin DH, Lee KH, Jeong LS, Yu J, Lee HW, Cho JY, Kim SY, Kim SW, Kim JB, Park KS, Chun YS. 2016. PPARγ neddylation essential for adipogenesis is a potential target for treating obesity. Cell Death Differ 23: 1296-1311.
-
-
15. Seale P, Conroe HM, Estall J, Kajimura S, Frontini A, Ishibashi J, Cohen P, Cinti S, Spiegelman BM. 2011. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J Clin Invest 121: 96-105.
-
16. Stanford KI, Middelbeek RJ, Townsend KL, An D, Nygaard EB, Hitchcox KM, Markan KR, Nakano K, Hirshman MF, Tseng YH, Goodyear LJ. 2012. Brown adipose tissue regulates glucose homeostasis and insulin sensitivity. J Clin Invest 123: 215-223.
-
17. Tine M, Kuhl H, Jastroch M, Reinhardt R. 2012. Genomic char- acterization of the European sea bass Dicentrarchus labrax reveals the presence of a novel uncoupling protein (UCP) gene family member in the teleost fish lineage. BMC Evol Biol 12: 1-15.
-
18. Villarroya F, Cereijo R, Villarroya J, Giralt M. 2017. Brown adipose tissue as a secretory organ. Nat Rev Endocrinol 13: 26-35.
-
19. Vishvanath L, Gupta RK. 2019. Contribution of adipogenesis to healthy adipose tissue expansion in obesity. J Clin Invest 129: 4022-4031.
-
20. Wang H, Liu L, Lin JZ, Aprahamian TR, Farmer SR. 2016. Browning of white adipose tissue with roscovitine induces a distinct population of UCP1+ adipocytes. Cell Metab 24: 835-847.
Copyright (C) 2024. The Korean Society of Marine Life Science