










JMLS 2019 December;4(2):63-69. http://jmls.or.kr Epub 2019 December 13
Copyright © 2019 by The Korean Society of Marine Life Science
Induced Ovulation by Hormone Treatments in Red Spotted Grouper, Epinephelus akaara
Jong-Youn Park; Department of Fisheries Science, Chonnam National University, Yeosu 59626, Korea
Jae-Kwon Cho; South East Fisheries Research Institute, National Institute of Fisheries Science, Tongyeong 53085, Korea
Chung-Kug Park; Jeonnam Maritime and Fisheries Science Museum, Yeosu 59771, Korea
Kyeong-Ho Han; Department of Fisheries Science, Chonnam National University, Yeosu 59626, Korea
Chang-Gi Hong; South Sea Fisheries Research Institute, National Institute of Fisheries Science, Yeosu 59780, Korea
- Abstract
본 연구는 붉바리 배란유도를 위해 다양한 호르몬과(ovaprim, pimozide, LHRHa, HCG)과 LHRHa의 농도별 효과를 조사하였다. LHRHa는 50 μgkg-1, 100 μgkg-1, 150 μgkg-1, 200 μgkg-1의 농도로 처리하였다. 호르몬은 등 근육에 주사하였으며, 조사 결과 LHRHa 단독, 그리고 LHRHa와 Pimozide를 혼합하여 투여했을 때 가장 효과적이었으며, 두 실험구 사이의 차이는 거의 없는 것으로 나타났다. 따라서 LHRHa를 단독으로 사용하는 것이 가장 효율적인 것으로 판단된다. 그리고 LHRHa를 다양한 농도로 처리하여 배란유도 효과를 조사한 결과 100 μgkg-1의 농도로 투여했을 때 가장 우수한 것으로 조사되었다.
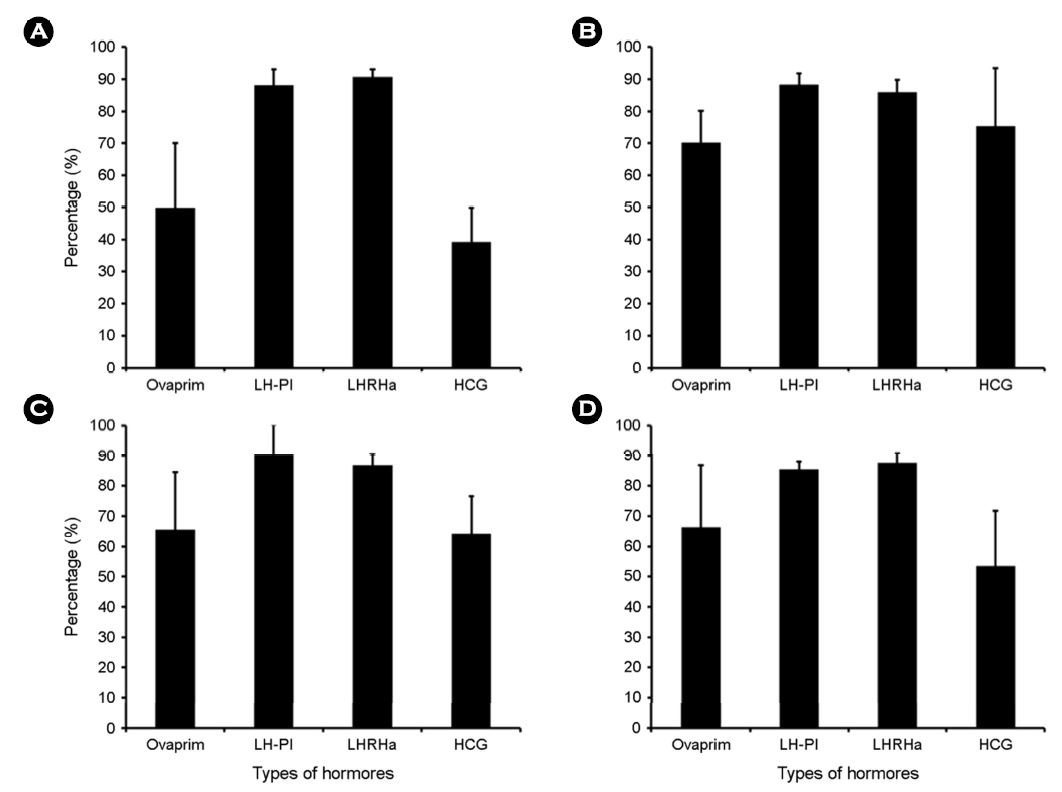
This study aims to investigate how ovulation induction of red spotted grouper, Epinephelus akaara varies when treated with different hormonal substances such as ovaprim, pimozide, LHRHa, and HCG. As for LHRHa, we injected it at four different concentrations: 50 μgkg-1, 100 μgkg-1, 150 μgkg-1, and 200 μgkg-1. All hormonal substances including LHRHa were injected into the back muscles. Buoyant, embryonic survival, fertilization, and hatching rate were the highest in LHRHa and LHRHa + pimozide group. When LHRHa was injected at different concentrations, buoyancy and fertilization rate were the highest in 100 μgkg-1 group. Embryonic survival and hatching rate were the highest in 150 μgkg-1. Based on these experimental results, we concluded that LHRHa at the concentration of 100 μgkg-1 is the most efficient for the ovulation induction in this fish species.
Keywords: Hormone(호르몬) Aquaculture(양식) Ovulation induction(배란유도) Grouper (바리과) Red spotted grouper(붉바리)
Correspondence to: Chang-Gi Hong; South Sea Fisheries Research Institute, National Institute of Fisheries Science, Yeosu 59780, Korea; E-mail : cknara@naver.com;
- Received
- 3 December 2019;
- Revised
- 4 December 2019;
- Accepted
- 9 December 2019.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
1. 실험구 및 실험어
바리과(Family Serranidae) 어류는 전 세계 3조원 이상의 시장규모를 가지고 있고, 중화권을 중심으로 고가로 소비되고 있는 어종으로, 아시아 지역에서 주로 양식하고 있다(Kohno et al., 1993; Harikrishnan et al., 2012). 그 중 붉바리, Epinephelus akaara는 중국, 대만, 동남아시아, 일본 중부이남 지역으로 대부분 아열대 및 열대 해역의 암초와 산호초가 있는 지역에 분포한다. 국내에서는 남해안 일부 연안, 특히 거문도 해역을 중심으로 남쪽으로는 제주도, 북쪽으로는 소리도 및 욕지도 등지에 분포하고 있다(Kohno et al., 1993; Harikrishnan et al., 2012). 현재까지 붉바리에 관한 연구로는 산란습성과 초기생활사(Ukawa and Higuchi, 1966), 생식소 발달(Hwang et al., 1988), 정자 동결보존(Qiutao et al., 2011) 등에 관한 연구가 보고되었다. 국내에서는 성장호르몬 발현(Kang et al., 2003), 성숙과 성전환(Lee et al., 1998), 먹이생물(Lee and Hur, 1998), 난질변화(Lee et al., 1997), 난 발생과 자치어 형태 발달(Park et al., 2016), 잡종 수정란 부화(Noh et al., 2015) 등 많은 연구가 보고되었다. 이러한 다양한 연구에도 불구하고 산업체에서 요구하고 있는 붉바리의 양질의 수정란 확보 기술과 초기먹이생물 등 종묘생산에 대한 연구는 미비한 실정이다.
산업적으로 중요한 어류의 종자생산에 있어 가장 중요한 요소 중 하나는 일시에 다량의 우량 수정란을 확보해야 하는 것이다(Hong et al., 2015). 자연산란이 어려운 어류의 수정란 생산을 위해서는 호르몬 처리를 통해 수정란 생산을 하고 있다(Chen, 1990; Tucker et al., 1994). 이러한 이유로 대부분 바리과 어류의 수정란 생산은 삽관법(cannulation)으로 난모세포의 내외의 크기를 확인한 후 호르몬을 처리하여 자연산란을 유도하거나 복부를 압박하여 채란, 채정하여 수정란을 생산한다. 국내에서도 붉바리 대량종자생산을 위해 어미를 해상가두리 또는 육상수조에서 관리하며 산란기에 어미를 선별하여 호르몬 처리 후 복부를 압박하여 수정란을 생산한다. 일반적으로 어류의 인위적인 배란유도를 위해 사용하고 있는 호르몬은 HCG (human chorionic gonadotropin), GnRH (gonadotropin releasing hormone), LHRHa (luteinizing hormone releasing hormone analogue) 등을 사용하는데, 최근에는 Pimozide를 혼합하여 사용하거나 Ovaprim을 이용하기도 한다(Watanabe et al., 1998; Tamaru et al., 1996; Hassin et al., 1997; Park et al., 1998; Marino et al., 2003). 이러한 호르몬 및 호르몬 유도체 처리에 의한 배란유도 방법은 같은 종에 있어서도 어미 사육관리의 환경조건 및 호르몬의 종류와 처리 농도에 따라 반응이 다르기 때문에 우량 수정란 확보를 위해서는 적정 호르몬의 종류와 처리 농도를 결정하는 것이 매우 중요하다(Soyano et al., 1993; Jang, 1996). 하지만 호르몬 종류 및 농도에 따른 효과에 대한 연구는 미비한 실정이다(Hong et al., 2015).
따라서 본 연구에서는 해상가두리 양식장에서 사육한 붉바리 어미의 안정적인 우량 수정란 확보를 위해서 Ovaprim, HCG, LHRHa 및 Pimozide를 처리하고 배란유도 효과와 호르몬 처리조건에 따른 수정률, 부상률, 난 발생생존율 및 부화율을 비교하였다.
재료 및 방법
1. 실험어
실험어는 2013년 6월에 전남 여수시 거문도 해역에서 채집한 자연산 붉바리를 해상가두리(5×5×5 m)에서 양성한 후 2014년 6월 성숙한 어미 후보군 중 복부팽만도와 생식공의 발달 정도에 따라 선별하여 2-phenoxyethanol (Sigma Co., USA)을 200 ppm으로 희석시킨 해수로 마취시켜 호르몬을 처리하였다.
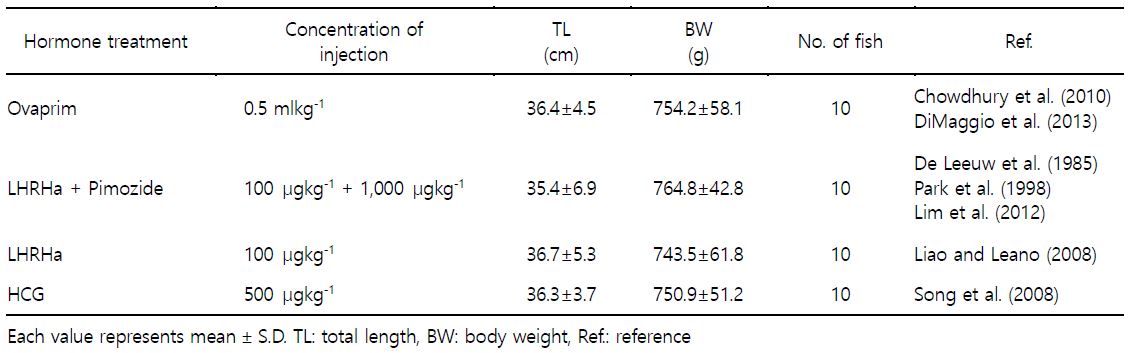
2. 호르몬 종류별 배란유도
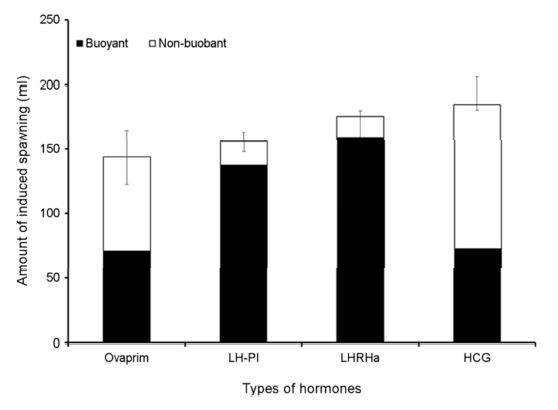
적정 호르몬 종류를 구명하기 위해 성숙한 암컷(전장 36.2± 5.1 cm, 체중 753.4±53.5 g) 각각 10마리씩 호르몬 종류별로 배란을 유도하였다(Table 1). 호르몬 종류는 Ovaprim [salmon GnRHa (sGnRHa: $\rm DArg^6$, $\rm Pro^{9}Net~sGnRH$), Domperidone; Syndel Laboratories LTD., Canada], Pimozide (Sigma Co. LLC., USA), LHRHa ($\rm desGly^{10}$, $\rm DAla^6$; Sigma Co. LLC., USA) 및 HCG (Daesung Microbiological Labs., LTD, Korea)였으며, 각 호르몬 농도는 Ovaprim는 0.5 $\rm mlkg^{-1}$, LHRHa + Pimozide은 100 $\rm \mu gkg^{-1}$ + 1,000 $\rm \mu gkg^{-1}$, LHRHa은 100 $\rm \mu gkg^{-1}$ 및 HCG은 500 $\rm IUkg^{-1}$였다. 각 종류별 호르몬을 제1극조 하부의 등 근육에 주사하고 48시간 경과 후 복부압박법으로 채란하여 건식법으로 수정시킨 후 부상률 및 수정률을 관찰하였고, 수정란을 대상으로 발생 과정에서 생존율 및 부화율을 계수하였다.
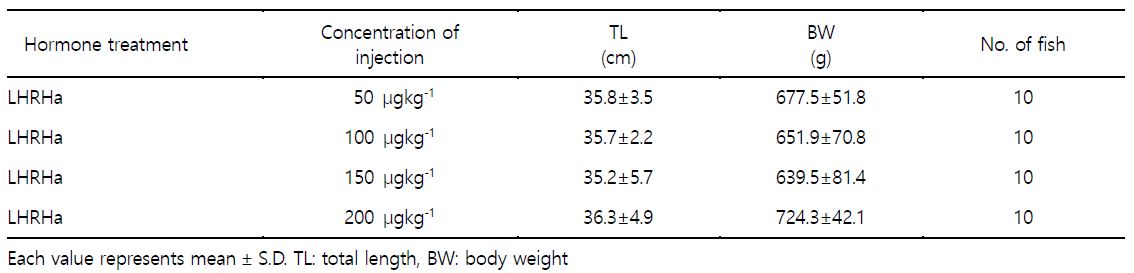
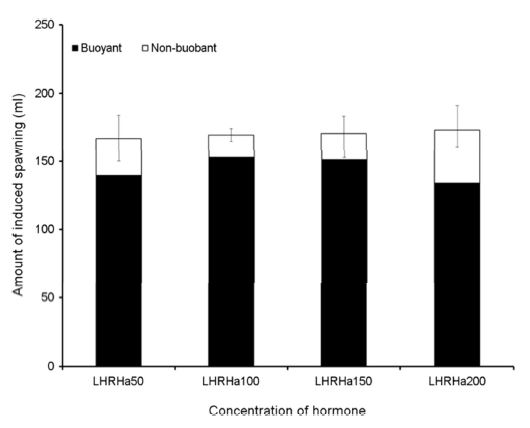
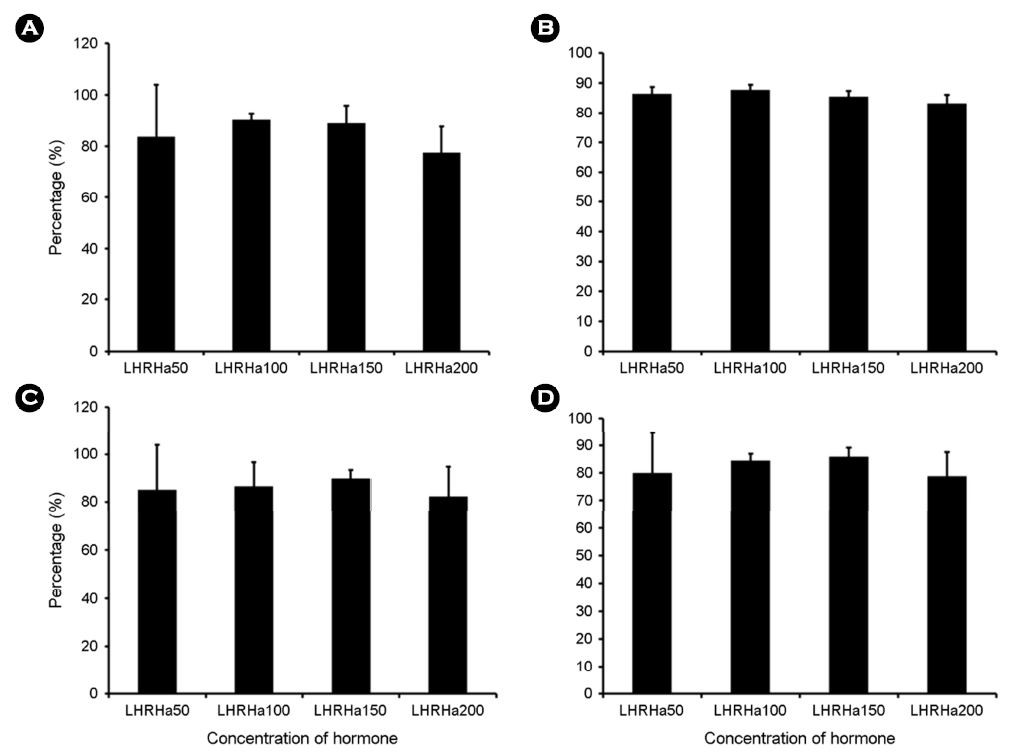
3. LHRHa 농도별 배란유도
호르몬 종류에 따른 배란유도 효과 결과를 토대로 LHRHa의 농도에 따른 배란유도 효과를 조사하였다. LHRHa의 처리 농도별 실험구는 50 $\rm \mu gkg^{-1}$, 100 $\rm \mu gkg^{-1}$, 150 $\rm \mu gkg^{-1}$ 및 200 $\rm \mu gkg^{-1}$ 구간으로 구분하여 제1극조 하부의 등 근육에 주사하고 48시간 경과 후 복부압박법으로 채란하여 건식법으로 수정시킨 후 부상률 및 수정률을 관찰하였고 수정란을 대상으로 발생과정에서 생존율 및 부화율을 측정하였다(Table 2). 각 실험구에서 암컷(전장 35.8±4.1 cm, 체중 673.3±61.5 g) 각각 10마리씩 호르몬 처리를 하였다.
- References
-

-
2. Chowdhury M, Chatterjee A, Mondal, A, Chatterji U. 2010. Ovaprim abrogates expression of GnRH receptor-II in the Indian catfish. International Journal of Biology 2.

-
3. Crim LW, Nestor JJ Jr, Wilson CE. 1988. Studies of the biological activity of LHRH analogue in the rainbow trout, landlocked salmon, and the winter flounder. Gen Comp Endocrinol 71: 372-382.
-
4. De Leeuw R, Goos HJ, Richter CJJ, Eding EH. 1985. Pimozide-LHRHa induced breeding of the african catfish, Clarias gariepinus (Burchell). Aquaculture 44: 295-302.
-
5. DiMaggio MA, Broach JS, Ohs CL. 2013. Evaluation of ovaprim and human chorionic gonadotropin doses on spawning induction and egg and larval quality of pinfish, Lagodon rhomboides. Aquaculture 414-415.
-
6. Donaldson EM, Hunter GA. 1983. Induced final maturation, ovulation and spermiation in cultured fishes. In: Hoar, W.S., Randall, D.J., Donaldson, E.M. (Eds.), Fish Physiology. Reproduction Vol IXB Academic press Orlando FL 351-403.
-
7. Harikrishnan R, Kim JS, Balasundaram C, Heo MS. 2012. Immunomodulatory effects of chitin and chitosan enriched diets in Epinephelus bruneus against Vibrio alginolyticus infection. Aquaculture 326-329.
-
8. Hassin SD, De Monbrison Y, Hanin A, Elizur Y, Zohar, Popper DM. 1997. Domestication of the white grouper, Epinephelus aeneus 1. Growth and reproduction. Aquaculture 156: 305 -316.
-
9. Hodson R, Sullivan CV. 1993. Induced maturation and spawning of domestic and wild striped bass, Morone saxatilis (Walbaum), broodstock with implanted GnRH analogue and injected HCG. Aquacult Fish Manage 24: 389-398.
-
10. Hong CG, Cho JK, Park JY, Son MH, Park JM, Han KH, Kang HW. 2015. Ovulation induction effect of sevenband grouper, Epinephelus septemfasciatus by treating hormones. J Kor Soc Fish Mar Sci 27: 981-989.
-
11. Hwang SI, Lee YD, Song CB, Rho S. 1988. Gonadal Development and the effects of 17α-methyltestosterone. J Aqua Korean 11: 173-182.
-
12. Jang SI. 1996. Induced ovulation by using human chorionic gonadotropin and gonadotropin-releasing hormone analogue plus pimozide in yellow puffer, Takifugu obscurus. Korean J Aquacult 9: 3-10.
-
13. Kang GY, Song CB, Lee JH. 2003. Cloning of growth hormone complementary DNA from red spotted grouper (Epinephelus akaara) and its expression in E.coli. J Aqua Korean 16: 110 -117.
-
14. Kohno H, Diani S, Supriatna A. 1993. Morphological development of larval and juvenile grouper, Epinephelus fuscoguttatus. Japanese Journal of Ichthyology 40: 307-316.
-
15. Kwon HC, Choi NJ, Park HY. 1996. Induced ovulation on catfish (Silurus asotus) by GnRH-Analogue. Korean J Aquacult 9: 205-213.
-
-
17. Lee CK, Hur SB, Park S, Kim BG. 1997. Qualities of spawned eggs during the spawning period in red spotted grouper, Epinephelus akaara. J of Aqua 10: 463-472.
-
18. Lee CK, Hur SB. 1998. Effect of live food and water temperature on larval survival of red spotted grouper, Epinephelus akaara. J of Aqua 11: 565-572.
-
19. Lee CK, Hur SB, Ko TS, Park S. 1998. Maturation, sex ratio and sex-reversal of red spotted grouper, Epinephelus akaara. J of Aqua 11: 573-580.
-
20. Lee CK, Hur SB. 1997. Yolk resorption, onset of feeding and survival potential of larvae of red spotted grouper, Epinephelus akaara. J of Aquacult 10: 473-483.
-
-
22. Lim SG, Kim KS, Kim YS, Han HG, Kim EO. 2012. Effect of HCG, LHRHa, Ovaprim and Pimozide on Ovulation Induction in Long Snout Bullhead Leiocassis longirotris. Dev Reprod 16: 137-143.
-
23. Lin H, Peng RC, Lu LZ, Zhou XJ, Kraak GVD, Peter RE. 1985. Induction of ovulation in the Loach (Paramisgurnus dabryanus) using pimozide and [D-ALA6, PRO9-N-Ethylamide]-LHRH. Aquaculture 46: 333-340.
-
24. Marino G, Panini E, Longobardi A, Mandich A, Finoia MG, Zohar Y, Mylonas CC. 2003. Induction of ovulation in captive-reared dusky grouper, Epinephelus marginatus (Lowe, 1834), with a sustained-release GnRHa implant. Aquaculture 219: 841-858.
-
25. Noh CH, Kim KS, Myoung JG, Cho JK, Yun NJ, Lim HG, Bang IC. 2015. The hatchability of fertilized eggs of interspecific hybrid between red spotted grouper (Epinephelus akaara) and brown-marbled grouper (E. fuscoguttatus). Kor J Ichthyol 27: 16-20.
-
26. Okumura S, Sakae K. 1993. Effect of LHRH-a cholesterol pellet on ovarian maturation and spawning in roundnose flounder, Eopsetta grigorjewi. Suisanzoshoku 41:13-18.
-
27. Park HY, Lee JY, Lee YJ, Kwon HC. 1998. Induction of ovulation by LHRH-a and pimozide in the bullhead, Pseudobagrus fulvidraco. Korean J Aquacult 11: 151-158.
-
28. Park JY, Cho JK, Son MH, Kim KM, Han KH, Park JM. 2016. Artificial spawning behavior and development of eggs, larvae and juveniles of the red spotted grouper, Epinephelus akaara in Korea. Dev Reprod 20: 31-40.
-
29. Qiutao HE, Gang LU, Kai C, Enhui Z, Qiongshan F, Hansheng W, Jing L, Changjiang H, Qiaoxiang D. 2011. Sperm cryopreservation of the endangered red spotted grouper, Epinephelus akaara, with a special emphasis on membrane lipids. Aqua- culture 318: 185-190.
-
30. Sherwood NM, Parker DB, McRory JE, Lescheid DW. 1994. Molecular evolution of growth hormone-releasing hormone and gonadotropin-releasing hormone. (in) Sherwood, N.M., Hew, C.L. (eds.), Fish Physioogy. Molecular Endocrinology of Fish Vol 8 Academic Press New York 3-66.
-
31. Song YB, Baek HJ, Kim HB, Soyano K, Kim SJ, Lee YD. 2008. Induction of maturation and ovulation with HCG treatment in the sevenband grouper Epinephelus septemfasciatus. Korean J Aquacult 21: 96-101.
-
32. Sokolowska M, Peter RE, Nahorniak CS, Pan CH, Chang JP, Crim LW, Weil C. 1984. Induction of ovulation in goldfish, Carassius auratus, by pimozide and analogues of LH-RH. Aquaculture 36: 71-83.
-
33. Soyano KT, Nagae SM, Yamauchi K. 1993. Effects of thyroid hormone on gonadotropin-induced steroid production in medaka, Oryzias latipes, ovarian follicles. Fish Physiology and Bio- chemistry 11: 265-272.
-
34. Tamaru CS, Carlstrom CT, Fitzgerald Jr. WJ, Ako H. 1996. Induced final maturation and spawning of the marbled grouper, Epinephelus microdon capture from spawning aggregations in the republic of Palau, Micronesia, J World Aqua Soc 27: 363-372.
-
35. Tucker Jr. JW, Parsons JE, Ebanks GC, Bush PG. 1994. Induced spawning of Nassau grouper, Epinephelus striatus. J World Aqua Soc 22: 187-191.
-
36. Ukawa M, Higuchi M. 1966. Spawning habits and early life history of a serranid fish, Epinephelus akaara. Japan J Ichthyol 13: 156-161.
-
37. Watanabe WO, Ellis EP, Ellis SC, Feeley MW. 1998. Progress in controlled maturation and spawning of summer flounder Paralichthys dentatus broodstock. J World Aqua Soc 29: 393 -404.
-
38. Zohar Y, Goren A, Tosky M, Pagelson G, Leibovitz D, Koch Y. 1989. The bioactivity of gonadotropin-releasing hormones and its regulation in the gilthead seabream, Sparusaurata: in vivo and in vitro studies. Fish Physiol Biochem 7: 59-67.
-
39. Zohar Y, Mylonas CC. 2001. Endocrine manipulations of spawning in cultured fish: from hormones to genes. Aquaculture 197: 99-136.
Copyright (C) 2025. The Korean Society of Marine Life Science