










JMLS 2018 December;3(2):51-58. http://jmls.or.kr Epub 2018 December 17
Copyright © 2018 by The Korean Society of Marine Life Science
Comparison of Hsp90 and CYP1A Expression Patterns by Water Temperature Stress in Atlantic Salmon (Salmo salar)
Han Seung Kang; Department of Genome Research, MS BioLab, Daejeon 34576, Korea
Jae-Hee Song; Tidal Flat Research Institute, National Fisheries Research & Development Institute, Kunsan 54014, Korea
Hee Woong Kang; Tidal Flat Research Institute, National Fisheries Research & Development Institute, Kunsan 54014, Korea
- Abstract
수온의 변화는 어류의 거의 모든 생리학적 부분에 영향을 미친다. 기후 변화로 인한 수온의 상승은 어류에게 물리적 피해를 줄 수 있다. 이 연구는 최적의 수온(15°C)보다 높은 수온(20°C)에서의 대서양 연어의 건강상태를 평가하기 위해 수행하였다. 간 조직은 열 적응에 중요한 대사 기능을 발휘하기에 본 연구에 간 조직을 사용하였다. 생체지표유전자의 개발을 위한 분석 방법으로는 NGS RNAseq 방법을 사용하였고, 생체지표유전자의 발현 양상을 관찰하기 위한 분석 방법으로는 RT-qPCR을 사용하였다. NGS RNAseq 분석을 통해 1,366개의 차별적 발현 유전자를 확인하였으며, 그 중에서 880개의 증가하는 유전자와 486개의 감소하는 유전자를 확인하였다. 생체지표유전자로는 heat shock protein 90 alpha (Hsp90α), heat shock protein 90 beta (Hsp90β) 및 cytochrome P450 1A (CYP1A)을 선정하였는데 이들 유전자는 NGS RNAseq 분석에서 수온의 변화에 민감하게 반응하는 유전자들이었다. 이들 유전자의 RT-qPCR을 통한 발현 양상은 NGS RNAseq 분석과 유사하게 나타났다. 이 연구의 결과는 다른 어종에도 적용할 수 있으며, 산업적으로도 유용하다고 생각된다.
Variations in water temperature are known to affect almost every part of fish physiology. The rise in water temperature due to climate change can physically damage fish. This study was conducted to evaluate the health status of the Atlantic salmon (Salmo salar) at high water temperature (20°C) than the optimum water temperature (15°C). Liver tissue exerts important metabolic functions in thermal adaptation. Therefore, liver tissue was used in this study. The evaluation method is to develop the biomarker gene using NGS RNAseq analysis and to examine the expression pattern using RT-qPCR analysis. The NGS RNAseq analysis revealed 1,366 differentially expressed genes, among which 880 genes were increase expressed and 486 genes were decrease expressed. The biomarker genes are such as heat shock protein 90 alpha (Hsp90α), heat shock protein 90 beta (Hsp90β) and cytochrome P450 1A (CYP1A). The selected genes are sensitive to changes in water temperature through NGS RNAseq analysis. Expression patterns of these genes through RT-qPCR were similar to those of NGS RNAseq analysis. The results of this study can be applied to other fish species and it is considered to be useful industrially.
Keywords: Salmo salar(대서양연어) Water temperature(수온) Biomarker gene(생체지표유전자) NGS RNAseq(차세대유전체분석법 RNAseq) RT-qPCR(역전사-실시간정량적PCR)
Correspondence to: Hee Woong Kang; Tidal Flat Research Institute, National Fisheries Research & Development Institute, Kunsan 54014, Korea; E-mail : hwgang@korea.kr;
- Received
- 4 December 2018;
- Revised
- 5 December 2018;
- Accepted
- 6 December 2018.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
수온, 산소 포화도 및 염분 농도 등은 해양생태계에 서식하는 생물들에게는 매우 중요한 환경요인으로 작용하고 있다. 이들 환경요인은 자연생태계 해양생물의 서식 및 삶의 전반과 더불어 양식 대상 해양생물에도 영향을 미쳐 성장, 생식, 대사 및 삼투압조절 등 해양생물의 생리적 변화에 영향을 미친다고 보고하였다(Chapple et al., 1998; Norris et al., 2010; Hvas et al., 2018; Pavlosky et al., 2019). 특히 환경요인 중에서 수온은 해양생물에게 매우 중요하다고 알려져 있다. 수온의 변화는 일반적으로 계절에 영향을 받는 것이 순리이나 이산화탄소 및 온실가스 배출 등에 의한 지구온난화가 기후변화의 원인으로 작용하여 수온의 변화를 야기하고 있다(Sampaio et al., 2018). 기후변화에 의해 발생할 수 있는 수온의 상승은 산소 용해도의 감소 및 산소를 운반하는 헤모글로빈의 결합력 약화를 통한 저산소증(hypoxia)을 유발하여 해양생물에게 영향을 줄 수 있다(Quinn et al., 2011a). 수온변화는 해양생물의 서식처 변경이나 회유경로 변화 등의 생태환경 및 해양생물의 체내 항상성에 교란을 일으켜 스트레스를 유발한다고 알려져 있다(Caissie, 2006; Crossin et al., 2008; Wang et al., 2015). 또한 어류에 있어서 수온의 변화는 생리작용의 거의 모든 부분에 영향을 미치는 것으로 알려져 있다(Somero, 2004). 연어에서도 수온의 변화는 배아의 생존 및 형질 형성 등 초기 발생 과정에 영향을 미치며 또한 개체의 이동, 은화(smoltification), 성장 및 생존에 영향을 미친다고 알려져 있다(McCormick et al., 1999; Swansburg et al., 2002; Elliott and Elliott, 2010; Somero, 2010; Burt et al., 2012; Hevrøy et al., 2012; Morita and Nakashima, 2015).
생물을 대상으로 환경요인의 변화에 의한 체내 생리학적인 변화를 통한 생물 개체의 건강상태 등의 평가는 개체를 대상으로 직접적인 조사를 통한 평가도 있지만, 발현 유전자 조사 및 유전자 발현 양상의 분석을 통해서도 빠르고 경제적으로 평가할 수 있다. 수온의 변화에 따른 고수온 및 저산소증 등과 관련한 유전자 발현 연구 등은 연어(Somero, 2004; Evans et al., 2011; Quinn et al., 2011a, 2011b), 저온 적응 남극 서식 어류(Thorne et al., 2010; Windisch et al., 2011) 및 어류(Kassahn et al., 2007; Healy et al., 2010; Logan and Somero, 2011; Liu et al., 2013) 등을 대상으로 수행하였으며, 연구 결과 열 충격 단백질(heat shock proteins) (Healy et al., 2010; Evans et al., 2011; Logan and Somero, 2011; Quinn et al., 2011a), 세포성장(cell growth) (Kassahn et al., 2007), 세포주기 및 세포사멸(cell cycle and apoptosis) (Logan and Somero, 2011), 염증반응(inflammatory response) (Thorne et al., 2010) 및 산화스트레스(oxidative stress) (Thorne et al., 2010) 관련 유전자군에서 차별적 유전자 발현이 관찰되었다고 보고하였다.
본 연구는 연어목(Salmoniformes), 연어과(Salmonidae) 어류인 대서양 연어(Atlantic salmon, Salmo salar)를 대상으로 수온변화로 인한 개체 스트레스 상태를 생체지표유전자(biomarker gene)의 발현 양상을 통해 평가하였다. 평가를 위한 생체지표유전자의 개발 및 선정은 차세대유전체분석법(NGS, Next Generation Sequencing) 중 RNAseq을 이용하였고, RT-qPCR 방법을 이용하여 유전자 발현 양상을 확인하였다. 생체지표유전자로 평가에 선정된 유전자는 heat shock protein 90 alpha (Hsp90α), heat shock protein 90 beta (Hsp90β) 및 cytochrome P450 1A (CYP1A) 유전자들이다. 선정된 유전자들은 수온의 변화에 민감하게 반응하여 수온 상승에 유의적으로 증가 및 감소한 유전자들로서 스트레스 및 대사에 관여하는 유전자들이다. 열 충격 단백질인 heat shock protein은 온도, 염분, 산화스트레스 및 중금속 등의 환경인자에 의해 영향을 받으면 유전자의 발현에 차이를 보이는 대표적인 스트레스 유전자로서 분자량을 기준으로 Hsp110, Hsp100, Hsp90, Hsp70, Hsp60 및 저 분자량 Hsp 등이 있다(Li et al., 2009; Colinet et al., 2010; Xu and Qin, 2012; Teixeira et al., 2013). 이 중에서 Hsp90은 세포분화 및 발생에 관여하며 다양한 생물에서 스트레스 반응 시 단백질조절, 세포사(apoptosis)조절 및 신호전달에 관여한다고 알려져 있다(Wu and Chu, 2008). Cytochrome P450 (CYP)은 heme을 포함하고 있는 효소로서 내인성 및 외인성 물질에 대한 제1상 약물대사반응의 대사에 관여한다. 이때 외인성 물질로는 약물, 발암물질 및 화학적 오염물질 등이 해당된다. 어류에서의 CYP1A는 환경오염물질로 분류되는 많은 화학물질에 의해 유도되는 성질로 인해 수중 환경오염의 biomarker로 활용도가 높다가 알려져 있다(Fent, 2001).
본 연구의 목적은 대서양 연어를 대상으로 수온변화에 따른 NGS RNAseq 연구 수행을 통해 개발된 유전자로서, 유전자의 기능에 따른 생체지표유전자의 개발을 목표로 연구진의 이전 면역관련 생체지표유전자의 보고(Kang et al., 2018)에 이어서 환경 스트레스 및 대사에 관여하는 유전자인 Hsp90α, Hsp90β 및 CYP1A를 이용하여 생체지표유전자로의 가능성 확인 및 스트레스 정도를 RT-qPCR 방법을 통해 평가하고자 하였다.
재료 및 방법
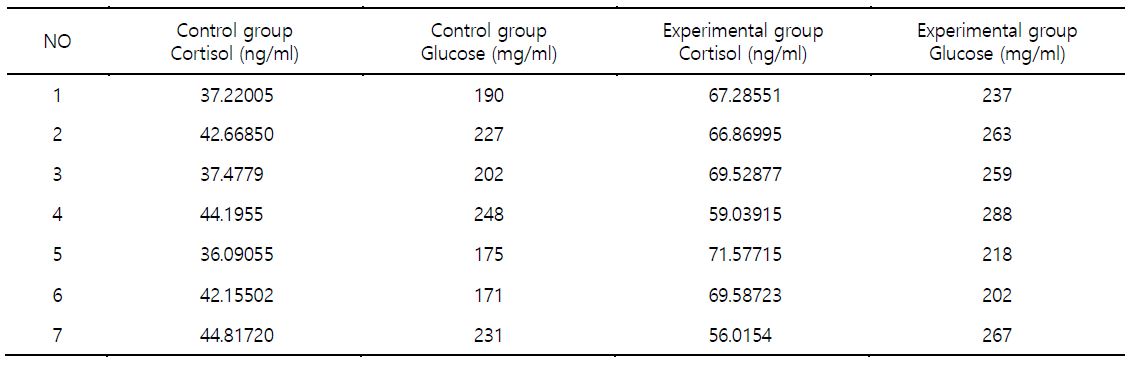
1. 실험동물
대서양 연어(11.9~13.5 cm)를 유수식 사육장에서 사육하며 실험하였다(Fig. 1). 수온에 따른 노출은 대조구(15℃) 및 실험구(20℃)에서 3시간 시행하였다. 노출 시간을 3시간으로 설정한 것은 수온의 장시간 노출은 초반에 민감한 반응을 나타내다 시간의 경과에 따라 적응이 되는 현상의 결과가 많이 보고되었다. 따라서 수온변화에 따른 생물의 초기반응에 관한 관심이 연구의 목적이다. 각 실험구별 연어를 대상으로 혈액성상 분석을 시행하여 스트레스 지표로 알려진 cortisol 및 glucose 농도를 측정하였다. 실험어의 선택 시 cortisol 및 glucose 분석 결과를 참조하여 대조구의 경우 농도의 수치가 낮은 것을 선정하였으며, 고수온 실험구에서는 cortisol 및 glucose 농도의 수치가 높은 것을 선정하였다. 실험에 사용한 연어는 각각 7마리로서 대조구 및 실험구에서 각각의 개체번호 1, 2, 3, 4, 5, 6 및 7번 개체를 선정하여 실험에 사용하였다(Table 1). 선정된 실험어에서 각각 간 조직을 수집한 후에 액체질소를 이용하여 동결 전 처리 후 -80℃ 초저온 냉동고에 보관하였다(Kang et al., 2018).

2. Total RNA 추출
Total RNA 추출은 GeneAll RiboEx kit를 사용하였다. 저온 동결 보존된 간 조직을 grinder로 마쇄한 후 RiboEx 용액(1 ml/100 mg)을 넣고 혼합한 후 실온에서 5분간 방치하였다. Chloroform 0.2 ml를 넣어주고 혼합 후 실온에서 2분간 방치하였다. 원심분리를 시행하여 상등액을 새로운 tube에 옮겨 담고 isopropyl alcohol 0.5 ml를 넣어 10분간 실온에서 방치하였다. 원심분리를 시행하여 pellet를 생성시킨 후, 상등액을 제거하고 75% EtOH로 세척한 후 DEPC-water로 녹여 -80℃ 초저온 냉동고에 보관하였다. RNA quality 확인은 Agilent사의 Bioanalyzer RiboPico 6000 chip을 이용하여 18S/28S 비율 및 RIN (RNA Integration Number)을 조사하였다. 대조구 및 실험구 각각의 개체에서 추출한 total RNA는 동량을 얻은 후, 합쳐서 실험에 사용하였다(Kang et al., 2018).
3. NGS RNAseq 분석 및 생체지표유전자 선정
대조구(15℃) 및 실험구(20℃)에서 채집한 간 조직에서 추출한 total RNA는 DNase 및 Ribo-zero rRNA remove kit를 이용하여 mRNA 및 non-coding RNA를 포함한 total RNA를 정제하였다. 정제된 RNA는 short read로 sequencing하기 위해 random하게 fragmentation 시킨 후, 역전사 과정을 통해 cDNA를 합성하였다. 합성한 cDNA fragment 양쪽 끝에 서로 다른 adapter를 붙인 후, ligation 시켰다. 다음으로 sequencing을 위하여 PCR 증폭을 통해 양을 증폭시킨 후, size selection 과정을 통해 200~400 bp의 insert size를 확보한 다음, paired-end sequencing으로 cDNA fragment의 양쪽 말단으로부터 read의 length만큼 sequencing 시행하였다. Sequencing을 통해 얻어진 raw reads의 quality control 분석을 진행하여 전체적인 read의 quality와 total bases, total reads, GC (%) 등 기본 통계치를 생산하였다. 분석 결과의 bias를 줄이기 위해 low-quality를 가지거나 adaptor sequence, contaminant DNA, PCR duplicates와 같은 artifacts을 제거하는 전처리 과정을 수행하였다. 전처리 과정을 거친 reads들을 대상으로 splice를 고려한 HISAT2 프로그램을 이용하여 reference genome에 mapping한 후, aligned reads를 생성시켰다. Reference 기반 aligned reads의 paired 정보를 이용하여 StringTie 프로그램을 통한 transcript 어셈블리를 진행하였다. 대조구 및 실험구 각각의 간 조직 transcript quantification을 통해 얻은 발현량을 transcript length 및 depth of coverage를 고려한 normalization 값으로 계산하였다. FPKM (Fragments Per Kilo- base of transcript per Million mapped reads) 값으로 within normalization을 진행하여 expression profile을 추출하였다. 발현값을 통계적인 가설검증을 통하여 차별 발현하는 유전자 또는 transcripts를 선별하여 생체지표유전자로 선정하였다(Kang et al., 2018).
4. 중합효소연쇄반응(RT-qPCR)
cDNA 합성을 위한 역전사 반응(Reverse Transcription, RT)은 iScript cDNA synthesis kit (Biorad Co.)를 이용하였다. Total RNA 1 μg, iScript 5×Master mix 4 μl, iScript reverse transcriptase 1 μl 및 DEPC-water을 넣어 최종 반응용액 20 μl을 맞추어 42℃에서 1시간 반응하여 cDNA를 합성하였다. 실시간 중합효소연쇄반응(realtime-qPCR)은 iQ SYBR Green Supermix kit (BioRad)를 이용하여 수행하였다. cDNA 1 μl, primer 각각 1 μl, iQ SYBR Green Supermix (2×) 10 μl 및 DEPC-water을 넣어 최종 반응용액 20 μl되게 맞춘 후에 real-time PCR machine (CFX96, Biorad)를 이용하여 증폭하고 형광량을 분석하였다. 유전자를 증폭시키기 위한 반응 조건은 95℃에서 3분간 유지, 이후 95℃에서 30초, 55℃에서 30초, 72℃에서 30초를 35 cycles를 반복하였으며, 마지막으로 72℃에서 5분간 유지하였다. Melting curve의 분석은 0.5℃ 간격으로 55℃에서부터 95℃까지 상승시켰다가, 이후 30℃에서 5분간 유지하였다. 상대적인 유전자 발현량의 결정은 2-∆∆Ct 방법(comparative Ct method)을 이용하여 유전자의 발현량을 분석하였다. 내재표준유전자로는 house keeping 유전자인 β-actin (ACTB)을 사용하여 발현량을 normalization 시켰다. 프라이머 염기서열은 다음과 같다(Table 2). 실시간 중합효소연쇄반응은 5회 반복 실험하였다.

5. 통계학적 분석
대조군과 실험군과의 유의성 검정은 Student's t-test로 비교하였으며, p가 0.01 이하인 것만 유의한 것으로 하였다.
결과 및 고찰
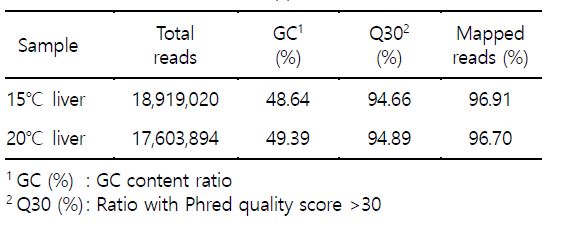
대서양 연어를 대상으로 NGS RNAseq 분석 연구의 목적은 수온의 변화에 보다 효율적이며, 민감하게 반응하는 전사체(transcriptome)를 다수 확보하고 그 중에서 생체지표유전자를 선정하고자 하는 것이다. 연어를 수온 15℃ 및 20℃에서 단기 3시간 노출 사육시킨 후 간 조직을 대상으로 추출한 total RNA를 이용하여 NGS RNAseq 분석을 수행한 결과로서 차별적 전사체를 다수 확보하였다. 수온 15℃ 사육 연어 간 조직에서 18,919,020개 reads가 생성되었으며, 총 길이의 합은 1.9G bp로 생산되었다. GC content (%)가 48.64%였으며, 염기품질점수 30 이상을 갖는 염기의 비율(Q30, %)은 94.66%이었고 전사체 mapping 효율은 96.91%였다. 수온 20℃ 사육 연어 간 조직에서는 17,603,894개 reads가 생성되었으며, 총 길이의 합은 1.8G bp로 생산되었다. GC content (%)가 49.39%였으며, 염기품질점수 30 이상을 갖는 염기의 비율(Q30, %)은 94.89%이었고 mapping 효율은 96.7%를 보였다(Table 3). 또한 |fc|>=2 조건을 만족하는 차별적 발현 유전자는 1,366개였고 그 중에서 증가 발현된 유전자 880개, 감소 발현된 유전자 486개였다(Fig. 2). 연구 결과 중에서 면역반응에 주로 관여하는 일부 유전자들에 대해서는 이전 보고(Kang et al., 2018)하였으며, 본 연구는 연장선상으로 스트레스 및 대사에 관여하는 유전자들의 연구 결과이다.

본 연구에서는 평가 대상 생체지표유전자로 ① Hsp90α, ② Hsp90β ③ CYP1A를 선정하였다(Table 4). 이들 유전자들은 NGS RNAseq 분석을 통해서 발현이 증가하거나 감소한 유전자들이다. 이들 유전자를 RT-qPCR 반응 분석을 통해 발현 양상을 살펴본 결과 fold change 값은 차이가 있었으나 유의적 증가 및 감소의 양상은 NGS RNAseq 결과와 유사하게 나타났다(Table 4, Fig. 3).
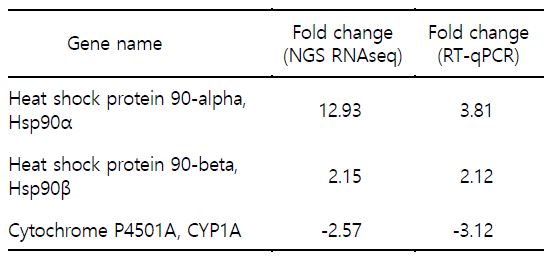

본 연구에서 발현 분석에 선정 및 이용한 유전자들의 기능은 잘 알려진 것도 있으나, 연어에서의 연구는 충분하지 않은 현실이다. 어류와 수온과 관련한 Hsp90 연구로는 다수의 연구 결과가 보고되었다. 무지개송어(Oncorhynchus mykiss)를 대상으로 수온과 Hsp90의 발현을 살펴본 연구에서는 간세포 배양의 실험 과정에서 배양온도를 높이게 되면 Hsp90 유전자 발현은 온도에 비례하여 발현이 높아졌다가 일정시간 경과 후 발현이 낮아지는 양상을 보였다(Sathiyaa et al., 2001). 그러나 열 충격 단백질인 Hsp가 생물종에 따라 다르게 발현되는 경우도 보고되었다. 송사리(Danio rerio)는 높은 수온의 스트레스를 주었을 경우 Hsp70의 발현은 초기에 높게 나타나다 시간의 경과에 따라 감소하는 경향을 나타냈으나, Hsp90의 발현은 수온변화에 차이가 나타나지 않았다(Murtha and Keller, 2003). 왕연어(Oncorhynchus tshawytscha)라 칭하는 chinook salmon의 경우 심장조직에서 Hsp90의 발현이 높게 나타났으며 수온이 높을수록 Hsp90의 발현은 높게 나타났다(Palmisano et al., 2000). 담수와 해수를 이동하는 연어에서 환경요인인 고 수온 및 고 삼투압 스트레스에 의한 Hsp90의 발현을 조사한 결과 고 수온 스트레스에는 Hsp90의 발현이 증가하였으나 고 삼투압 스트레스에는 반응하지 않았다(Pan et al., 2000). NGS RNAseq을 통해 나타난 Hsp90α와 Hsp90β는 고 수온의 스트레스에 발현이 증가하였으며 RT-qPCR을 이용한 발현 분석에서도 증가한 양상을 보였다. 특히 Hsp90α가 Hsp90β에 비교하여 민감하게 발현하는 양상을 보여주었다. 연구 결과는 대서양 연어와 수온 스트레스 및 Hsp90 유전자와의 관련성을 보이는 기초 연구 결과로 중요하다고 판단된다. Cytochrome P 450 (CYP)는 척추동물, 무척추동물, 식물, 곰팡이, 세균, 효모 등에서 발견되며 포유류에서는 수 백여개의 CYP가 발견되어 연구 중이다. 아과(subfamily)로는 CYP1A와 CYP2B로 나누어지며 CYP1A는 대부분의 척추동물에서 발견되며, CYP2B는 어류, 양서류, 파충류 등에서는 발견되지 않는다(Stegeman and Hahn 1994). 어류에서 CYP1A 관련 연구는 화학물질 혹은 오염물질과의 연구가 대부분이다(Fent, 2001). 어류가 서식하는 수중생태계의 수온은 극지방이나 열대지방과 같이 폭넓은 범위이다. 어류들은 급격한 수온변화를 피하기 위해 그들 만의 서식범위에서 살지만 홍수나 태풍 등의 불가항력적인 자연재해에 따른 수온의 변화는 피할 수 없다. 이러한 수온의 변화에 따른 저온환경에 노출된 어류는 CYP1A가 증가한다는 보고가 있다(Anderson and Forlin, 1992). 본 연구에서 NGS RNAseq을 통해 나타난 CYP1A는 고 수온에서 감소하는 경향을 보였다. 또한 RT-qPCR 발현 분석에서도 감소한 양상을 보였다. 연어를 포함한 어류에서 외인성 물질이 아닌 환경요인인 수온과 CYP1A의 연구는 많지않은 현실에서 수온의 변화와 CYP1A 유전자와의 관련성을 보이는 기초 연구 결과로 중요하다고 판단된다.
본 연구에서 생체지표유전자로 선정한 유전자들의 주요 기능은 스트레스 및 대사 관련 유전자로서 수온 변화 등의 외부 환경요인에 민감하게 반응하여 발현하였다. 변화의 폭이 크지 않은 환경요인의 변화에도 해양에 서식하는 양식생물을 포함하는 해양생물에게는 항상성에 영향을 미쳐 폐사를 유도할 수 있다. 따라서 생물의 스트레스에 따른 건강상태의 주기적인 점검은 매우 중요하다. 생체지표유전자의 발현 분석을 통한 건강도의 평가는 저비용 고효율의 방법이다. 생체지표유전자의 발현을 통한 건강도의 평가는 사람을 대상으로 하는 진단의학에도 적용하여 질병 및 향후 발생 가능한 질병의 예측에 이용되고 있다. 생체지표유전자는 생물 개체의 건강도 평가에 주요 적용 가능하나 더불어 환경요인에 대비한 선제적 적응 대책의 수립, 지역별 서식환경과 양식생물 간의 관계에 대한 변화체계 모니터링 및 데이터베이스 구축을 통한 안정적인 양식생산성 예측을 위한 기반 확보로도 주요하게 사용 가능하다고 생각된다.
본 연구를 통한 결과는 다른 어종에도 적용 가능하기에 산업적으로 매우 유용하리라 판단되며, 학문적으로도 유용하게 적용되리라 생각된다.
- References
-
1. Anderson T, Forlin TL. 1992. Regulation of the cytochrome P450 enzymes system in fish. Aquat Toxicol 24: 1-20.

-
2. Burt JM, Hinch SG, Patterson DA. 2012. Parental identity influences progeny responses to incubation thermal stress in sockeye salmon Onchorhynchus nerka. J Fish Biol 80: 444-462.

-
-
4. Chapple JP, Smerdon GR, Berry RJ, Hawkins AJS. 1998. Seasonal changes in stress-70 protein levels reflect thermal tolerance in the marine bivalve Mytilus edulis L. J Exp Mar Biol Ecol 229: 53-68.
-
5. Colinet H, Lee SF, Hoffmann A. 2010. Temporal expression of heat shock genes during cold stress and recovery from chill coma in adult Drosophila melanogaster. FEBS J 277: 174-185.
-
6. Crossin GT, Hinch SG, Cooke SJ, Welsh DW, Patterson DA, Jones SRM, Lotto AG, Leggatt RA, Mathes MT, Shrimpton JM, Van Der Kraak G, Farrell AP. 2008. Exposure to high temperature influences the behaviour, physiology, and survival of sockeye salmon during spawning migration. Can J Zool 86: 127-140.
-
7. Elliott JM, Elliott JA. 2010. Temperature requirements of Atlantic salmon Salmo salar, brown trout Salmo trutta and Arctic charr Salvelinus alpinus: predicting the effects of climate change. J Fish Biol 77: 1793-1817.
-
8. Evans TG, Hammill E, Kaukinen K, Schulze AD, Patterson DA, English KK, Curtis JMR, Miller KM. 2011. Transcriptomics of environmental acclimatization and survival in wild adult Pacific soc- keye salmon (Oncorhynchus nerka) during spawning migra- tion. Mol Ecol 20: 4472-4489.
-
9. Fent K. 2001. Fish cell lines as versatile tools in ecotoxicology: assessment of cytotoxicity, cytochrome P450 1A induction potential and estrogenic activity of chemicals and environmental samples. Toxicol In Vitro 15: 477-488.
-
10. Healy TM, Tymchuk WE, Osborne EJ, Schulte PM. 2010. Heat shock response of killifish (Fundulus heteroclitus): candidate gene and heterologous microarray approaches. Physiol Genomics 41: 171-184.
-
11. Hevrøy EM, Waagbo R, Torstensen BE, Takle H, Stubhaug I, Jorgensen SM, Torgersen T, Tvenning L, Susort S, Breck O, Hansen T. 2012. Ghrelin is involved in voluntary anorexia in Atlantic salmon raised at elevated sea temperatures. Gen Comp Endocrinol 175: 118-134.
-
12. Hvas M, Folkedal O, Imsland A, Oppedal F. 2018. Metabolic rates, swimming capabilities, thermal niche and stress response of the lumpfish, Cyclopterus lumpus. Biol Open 7: bio036079 (1-9).
-
13. Kang HW, Kim KI, Lim HJ, Kang HS. 2018. Effect of water temperature on the expression of stress related genes in Atlantic salmon (Salmo salar) Fry. Korean J Environ Biol 36: 131-139.
-
14. Kassahn KS, Caley MJ, Ward AC, Connolly AR, Stone G, Crozier RH. 2007. Heterologous microarray experiments used to identify the early gene response to heat stress in a coral reef fish. Mol Ecol 16: 1749-1763.
-
15. Li P, Zha J, Zhou KY. 2009. Molecular cloning, mRNA expression, and characterization of HSP90 gene from Chinese mitten crab Eriocheir japonica sinensis. Comp Biochem Physiol B 153: 229-235.
-
16. Liu SK, Wang XL, Sun FY, Zhang JR, Feng JB, Liu H, Rajendran KV, Sun LY, Zhang Y, Jiang YL, Peatman E, Kaltenboeck L, Kucuktas H, Liu ZJ. 2013. RNA-Seq reveals expression signatures of genes involved in oxygen transport, protein synthesis, folding, and degradation in response to heat stress in catfish. Physiol Genomics 45: 462-476.
-
17. Logan CA, Somero GN. 2011. Effects of thermal acclimation on transcriptional responses to acute heat stress in the eurythermal fish Gillichthys mirabilis (Cooper). Am. J. Physiol-Regul Integr Comp Physiol 300: R1373-R1383.
-
18. McCormick SD, Cunjak RA, Dempson B, O'Dea MF, Carey JB. 1999. Temperature-related loss of smolt characteristics in Atlantic salmon (Salmo salar) in the wild. Can J Fish Aquat Sci 56: 1649-1658.
-
19. Morita K, Nakashima A. 2015. Temperature seasonality during fry out-migration influences the survival of hatchery-reared chum salmon Oncorhynchus keta. J Fish Biol 87: 1111-1117.
-
20. Murtha JM, Keller ET. 2003. Characterization of the heat shock response in mature zebrafish (Danio rerio). Exp Gerontol 38: 683-691.
-
21. Norris AL, Houser DS, Crocker DE. 2010. Environment and activity affect skin temperature in breeding adult male elephant seals (Mirounga angustirostris). J Exp Biol 213: 4205-4212.
-
22. Palmisano AN, Winton JR, Dickhoff WW. 2000. Tissue specific induction of hsp90 mRNA and plasma cortisol response in chinook salmon following heat shock, seawater challenge, and handling challenge. Mar Biotechnol 2: 329-338.
-
23. Pan F, Zarate JM, Tremblay GC, Bradley TM. 2000. Cloning and characterization of salmon hsp90 cDNA: upregulation by thermal and hyperosmotic stress. J Exp Zool 287: 199-212.
-
24. Pavlosky KK, Yamaguchi Y, Lerner DT, Seale AP. 2019. The effects of transfer from steady-state to tidally-changing salinities on plasma and branchial osmoregulatory variables in adult Mozambique tilapia. Comp Biochem Physiol A Mol Integr Physiol 227: 134-145.
-
25. Quinn NL, McGowan CR, Cooper GA, Koop BF, Davidson WS. 2011a. Identification of genes associated with heat tolerance in Arctic charr exposed to acute thermal stress. Physiol Genomics 43: 685-696.
-
26. Quinn NL, McGowan CR, Cooper GA, Koop BF, Davidson WS. 2011b. Ribosomal genes and heat shock proteins as putative markers for chronic, sublethal heat stress in Arctic charr: applications for aquaculture and wild fish. Physiol Genomics 43: 1056-1064.
-
27. Sampaio E, Lopes AR, Francisco S, Paula JR, Pimentel M, Maulvault AL, Repolho T, Grilo TF, Pousão-Ferreira P, Marques A, Rosa R. 2018. Ocean acidification dampens physiological stress response to warming and contamination in a commercially-important fish (Argyrosomus regius). Sci Total Environ 618: 388-398.
-
28. Sathiyaa R, Campbell T, Vijayan MM. 2001. Cortisol modulates hsp90 mRNA expression in primary cultures of trout hepatocytes. Comp Biochem Physiol B, Biochem Mol Biol 129: 679 -685.
-
29. Somero GN. 2004. Adaptation of enzymes to temperature: searching for basic "strategies". Comp Biochem Physiol B-Biochem Mol Biol 139: 321-333.
-
30. Somero GN. 2010. The physiology of climate change: how potentials for acclimatization and genetic adaptation will determine 'winners' and 'losers'. J Exp Biol 213: 912-920.
-
31. Stegeman JJ, Hahn ME. 1994. Biochemistry and molecular biology of monooxygenase: Current perspectives on forms, functions and regulation of cytochrome P450 in aquatic species. Pp. 87-204. In Aquatic toxicology, molecular, biochemical and cellular perspectives (Malins DM and GK Ostrander, eds). Lewis Publishers. Boca Raton.
-
32. Swansburg E, Chaput G, Moore D, Caissie D, El-Jabi N. 2002. Size variability of juvenile Atlantic salmon: links to environmental conditions. J Fish Biol 61: 661-683.
-
33. Teixeira T, Diniz M, Calado R, Rosa R. 2013. Coral physiological adaptations to air exposure: Heat shock and oxidative stress responses in Veretillum cynomorium. Journal of Experimental Marine Biology and Ecology 439: 35-41.
-
34. Thorne MAS, Burns G, Fraser KPP, Hillyard G, Clark MS. 2010. Transcription profiling of acute temperature tress in the Antarctic plunderfish Harpagifer antarcticus. Mar Genomics 3: 35-44.
-
35. Wang Y, Li L, Hu M, Lu W. 2015. Physiological energetics of the thick shell mussel Mytilus coruscus exposed to seawater acidification and thermal stress. Sci Total Environ 514: 261 -272.
-
36. Windisch HS, Kathover R, Portner HO, Frickenhau S, Lucassen M. 2011. Thermal acclimation in Antarctic fish: transcriptomic profiling of metabolic pathways. Am J Physiol-Regul Integr Comp Physiol 301: R1453-R1466.
-
37. Wu LT, Chu KH. 2008. Characterization of heat shock protein 90 in the shrimp Metapenaeus ensis: evidence for its role in the regulation of vitellogenin synthesis. Mol Reprod Dev 75: 952 -959.
-
38. Xu Q, Qin Y. 2012. Molecular cloning of heat shock protein 60 (PtHSP60) from Portunus trituberculatus and its expression response to salinity stress. Cell Stress Chaperones 17: 589 -601.
Copyright (C) 2024. The Korean Society of Marine Life Science