










JMLS 2018 June;3(1):9-21. http://jmls.or.kr Epub 2018 June 14
Copyright © 2018 by The Korean Society of Marine Life Science
Cytogenetic Characteristics of Cyprinidae between Diploid and Spontaneous Triploid in Major River of Korea
In Bon Goo; Inland Aquaculture Research Center, National Institute of Fisheries Science (NIFS), Jinhae 51688, Korea
Sang Gu Lim; Aquafeed Research Center, NIFS, Pohang 37517, Korea
Hyun Woo Gil; Bio-Monitoring Center, Sejong 30121, Korea
In-Seok Park; Division of Marine Bioscience, College of Ocean Science and Technology, Korea Maritime and Ocean University, Busan 49112, Korea
Cheol Young Choi; Division of Marine Bioscience, College of Ocean Science and Technology, Korea Maritime and Ocean University, Busan 49112, Korea
- Abstract
This study investigated cytogenetic and hematological and histological characteristics between diploid and spontaneous triploid on Cyprinidae (Crucian carp, Carassius auratus; crucian carp, C. cuvieri and common carp, Cyprinus carpio) in four major rivers of Korea. In our results, DNA contents of triploid Cyprinidae were 50% more than those of diploid Cyprinidae. Also, erythrocyte size of triploid Cyprinidae was 50% larger than those of diploid Cyprinidae. In all sampling rivers, sex ratios of C. auratus were biased to female, and especially, triploid groups of C. auratus were all female groups (p<0.05). In principle, sex ratios of C. cuvieri and common carp were equivalent between male and female.
Keywords: Carassius auratus C. cuvieri Cyprinus carpio Cytogenetic characteristics Spontaneous triploid
Correspondence to: In-Seok Park; Division of Marine Bioscience, College of Ocean Science and Technology, Korea Maritime and Ocean University, Busan 49112, Korea; E-mail : ispark@kmou.ac.kr;
- Received
- 5 February 2018;
- Revised
- 9 May 2018;
- Accepted
- 10 May 2018.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: English,
Full Text:

Introduction
Among members of the Cyprinidae family, hybridization is more widespread than in any other group of freshwater fish, which has resulted in formation of many natural and artificial hybrids (Kim, 1997; Yoon and Park, 2006). Crucian carp, Carassius auratus and common carp, Cyprinus carpio are both well-known species of freshwater fish in Europe (Kucinski et al., 2015). In addition, C. auratus, crucian carp, C. cuvieri and common carp, Cyprinus carpio are wide-spread freshwater s pecies in Korea (Fig. 1; Kim, 1997; Yoon and Park, 2006). Under the natural ecosystem, Cyprinidae (C. auratus, C. cuvieri and common carp) is in entire lakes, marshes and rivers in the Korean peninsula as well as in the several areas in Japan, China, Taiwan, Siberia and the European continent. In particular, one species of crucian carp (C. auratus) is an economically important aquacultural species belonging to the family Cyprinidae. The common name, crucian carp in Korea was identified, C. auratus, by means of morphology and electrophoretic analysis (Nam et al., 1989). The C. auratus can be categorized as a species complex because is problematical and due to genetic variation or morphological differences resulting from environmental influences (Nam et al., 1989).
Another species of C. cuvieri is a native species of Yodogawa river and the Biwakko lake region on the west coast of Japan (Yoon and Park, 2006). This fish species has been successfully introduced into the many waters in Korea in the 1970's (Yoon and Park, 2006). C. cuvieri is ranked at the highest among the fresh-water fishes in Korea as a game fish attracting millions of anglers owing to the quake of fingertips (Yoon and Park, 2006). Common carp is the most commercially important fish species in Korea. Common carp inhabited in fresh water with temperature range between 3 and 35℃ exhibiting tolerance to a wide variety of conditions (Kim and Kim, 2009).
Cyprinidae have been extensively studied genetically when compared with other fish groups (Labat et al., 1983; Yoon and Park, 2006). Especially, C. auratus and common carp were detected spontaneous triploid and tetraploid by cytogenetic analysis (Al-Sabti et al., 1983; Anjum and Jankun, 1994; Kim et al., 2002). Spontaneous triploid groups of C. auratus in Korea were all-female group and having specific spawning mechanism, gynogenesis and have same genetic characteristic with maternal line (Kim et al., 2002). Spontaneous triploid groups of common carp were not investigated about specific spawning mechanism and C. cuvieri was not detected spontaneous triploid until now.
There are numerous studies in the literature which have investigated various aspects of induced triploid fish identification methodology including analysis of chromosome sets, the microfluorimetry of nuclear DNA content, the nuclear DNA content by flow cytometry, the measurement of erythrocyte and nuclear size, the distinction of nucleolar number, the measurement of cell number, and the measurement of cell and nuclear size in different tissues (Seol et al., 2008). Flow cytometry has a wide variety of clinical applications in oncology for understanding surface expression, intracellular signaling, cell cycle content analysis, and a number of other interesting parameters (Vanparys et al., 2006). Recent advances in instrument platforms, calibration methods, and reagent quality have now made flowcytometry a promising tool for DNA content analysis (Estevam et al., 2011). These calibration packages can detect if the parameters are within acceptable ranges and thus allow for consistent sample acquisition over time. One of the advantages of flow cytometry is the rapidity of the measurement, making it possible to measure thousands of cells over a short period of time, and the ability for multi-color immunophenotyping (Estevam et al., 2011).
Numerous studies have demonstrated that erythrocyte cellular and nuclear dimensions are increased, and numbers of erythrocytes are decreased in triploids (Benfey, 1999; Seol et al., 2008). Therefore, it is easy to distinguish between diploid and triploid fish by assessing the size and number of erythrocytes, which are reduced in triploidy in proportion to the erythrocyte size (Benfey, 1999). In sweetfish, Plecoglossus altivelis, triploid specimens had larger erythrocytes and lower erythrocytes number than diploid specimens, and also showed higher hematological parameters (mean corpuscular volume and mean content of haemoglobin) and oxygen consumption was higher in triploid than in diploid (Aliah et al., 1991).
So, this study investigated DNA content, gonado somatic index, erythrocyte size and histological differences of gonad between diploid and spontaneous triploid in Cyprinidae in four major rivers of Korea.

Fig. 1. External morphology of crucian carp, Carassius auratus (a), crucian carp C. cuvieri (b) and common carp, Cyprinus carpio (c) used in this experiment. Scale bars are 5 cm.
Materials and Methods
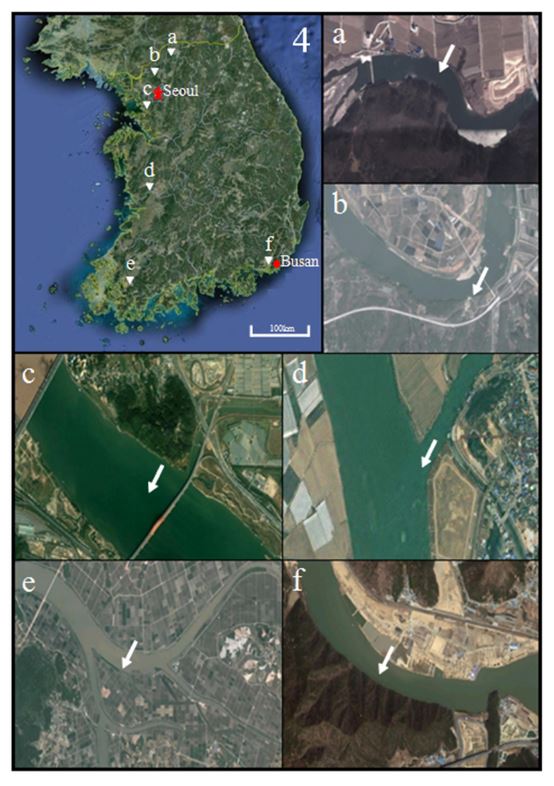
On June 2012, samples of crucian carp, Carassius auratus, crucian carp, C. cuvieri and common carp, Cyprinus carpio were trapped in the Han River, Hantan River, Imjin River, Kum River, Yongsan River and Nakdong River in Korea (Fig. 2). For hematological observation, blood samples were collected from caudal vein, and kept at 4℃ in heparin-treated polyethylene vials (70 IU/mℓ blood). Collected samples fixed in 10% neutral formalin solution (100 ml formalin, 6.5 g Na2HPO4 · 12H2O, 4.5 g KH2PO4, 900 ml distilled water), and 10% neutral formalin solutions of each sample were exchanged after 24 hours.
Ploidy identification was performed using flow cytometry. Ventral fins were collected from each fish, and fixed fins in 70% ethanol. The samples were analyzed using flow cytometry measurement. For flow cytometric analysis, tissue of ventral fin homogenized, filtered using 30 μm filter after centrifugation (Centrifuge Micro 17R, Hanil Science Industrial Co., Ltd, Incheon, Republic of Korea; 1,000 rpm, 10 min), removed supernatant liquid and added 0.5 ml CyStain DNA 2 step nuclei extraction buffer (CyStain DNA 2 step high resolution DNA staining kit, Partec, Germany) and 2 ml CyStain DNA 2 step staining buffer (CyStain DNA 2 step high resolution DNA staining kit, Partec, Germany). The red blood cells of mud loach, Musgurnus mizolepis were used as a standard reference (Seol et al., 2008). Identified samples and blood samples were separated with sampling site, sex and ploidy.
For determine sex ratio and calculate gonado somatic index (GSI), the gonads were enucleated from fixed sample of each group. GSI of each sample were calculated by following formula. GSI = (gonado weight / (total weight-gonado weight)) × 100. Twenty samples of each group used for histological observations. Extracted gonads were dehydrated via a standard ethanol series to 100%, cleared in xylene, embedded in paraffin. Six μm sections were deparaffinized and stained by Mayer's Hematoxylins & Eosin. Observation and evaluation was conducted via light microscopy (Carl Zeiss, Germany).
Erythrocytes and erythrocyte nuclei from stored blood samples were determined from dry blood smears. Air-dried blood smears were fixed in methyl alcohol and stained with May-Grünwald-Giemsa. Erythrocytes and their nuclear major and minor axis were determined in both live and fixed cells using a light microscope (Carl Zeiss, Germany) equipped with Axioskop 4.1 image analysis system. One-hundred and twenty cells were measured for each specimen. Surface areas and volume of erythrocyte cell and nuclear were calculated by follow equation:
**수식
where a and b are the major and the minor axis of the cell and of the nucleus, respectively (Sezaki et al., 1988). The cell and nuclear major and minor axis and surface for the cell and nucleus of specimens with each ploidy were compared.
The significant differences of sex ratio among experimental groups were assessed by χ2-test (p<0.05). One-way ANOVA and Duncan's multiple range test (Duncan, 1955) were utilized in order to assess the significance of the difference among the means of experimental groups, using SPSS version 9.0 (p<0.05; SPSS Int., USA).
Results
We noted significant differences of DNA content and spontaneous generation rate between sampling sites and species (Table 1). As shown in Table 1 and Fig. 3, DNA contents of crucian carp, Carassius auratus were similar to those of common carp, Cyprinus carpio (p>0.05). DNA contents of crucian carp, C. cuvieri were higher than those of the other species (p<0.05). DNA contents of triploid were half of diploid DNA contents in C. auratus, C. cuvieri and common carp (Fig. 3; p<0.05). DNA contents of Nakdong river in diploid of C. auratus were higher than those of the other sampling site, and the highest DNA contents were found in diploid of C. cuvieri and common carp in Han river (p<0.05). In triploid, the highest DNA contents were in C. auratus and C. cuvieri in Youngsan river (p<0.05), and DNA contents in common carp were similar in all sampling sites (p>0.05). The highest triploid spontaneous generation rate in C. auratus was in Hantan river, and triploid spontaneous generation rate of Han river in C. cuvieri was higher than in the other sampling sites (p<0.05). Triploid spontaneous generation rates of common carp showed no significant difference among all sampling sites (p>0.05).
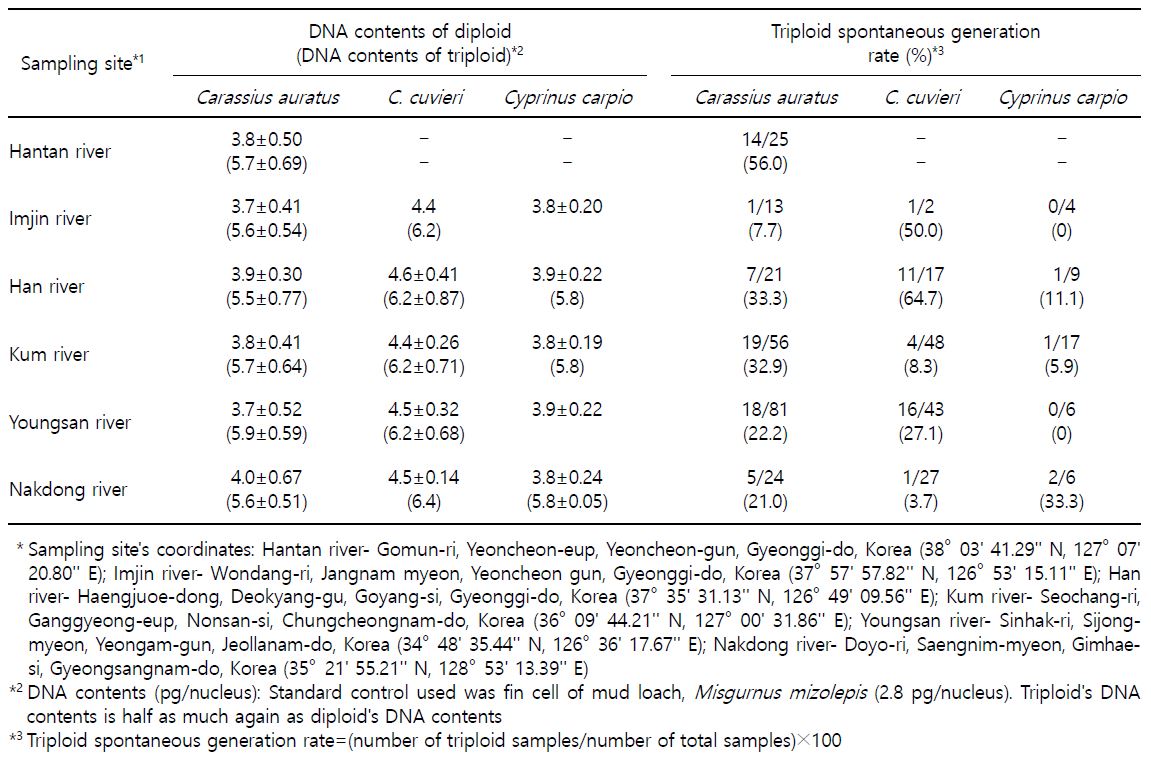
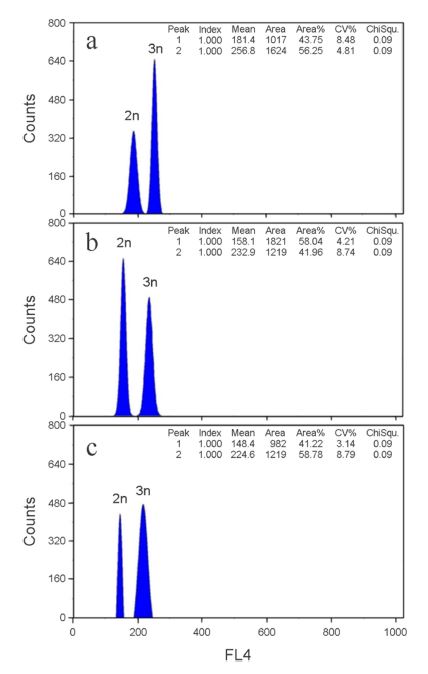
Table 2 shows the sex ratio and results of χ2-test (p<0.05). In C. auratus, sex ratios of diploid and triploid were significantly different in all sampling sites (p<0.05). Sex ratios of diploid C. auratus were biased to female, and especially, triploid groups of C. auratus were all female groups in all sampling site (p<0.05). In C. cuvieri, sex ratios of diploid were equivalent between male and female in Imjin river and Han river (p>0.05). Sex ratio of Kum river was biased to female, and sex ratios of Youngsan river and Nakdong river were biased to male (p<0.05). Sex ratios of triploid were equivalent between male and female in all sampling site (p>0.05). In common carp, sex ratio of diploid were equivalent between male and female, and sex ratios of triploid were no significant difference (p>0.05).

In all species, differences of the gonad size between diploid and triploid were determined by naked eye (Fig. 4). As shown in Fig. 4, testis and ovarium of triploid C. auratus and C. cuvieri were larger than those of diploid C. auratus and C. cuvieri, and ovarium of common carp was larger than ovarium of diploid common carp. However, testis of diploid common carp was smaller than testis of triploid common carp. Table 3 shows the difference of gonado somatic index (GSI) between sex and ploidy in C. auratus, C. cuvieri and common carp. GSI of triploid were half as much again as those of diploid in female of C. auratus, C. cuvieri and male of common carp (p<0.05). However, GSI of diploid in female of common carp were higher than those of triploid. In male and female, GSI of all diploid species were no significant difference among all sampling site, also, GSI of all triploid species were no significant difference among all sampling site (p>0.05). GSI of diploid male and diploid female in common carp were highest than those in C. auratus and C. cuvieri (p<0.05). GSI of triploid male in common carp were highest than those in C. cuvieri, and GSI of triploid female in common carp were highest than those in the other species (p<0.05).


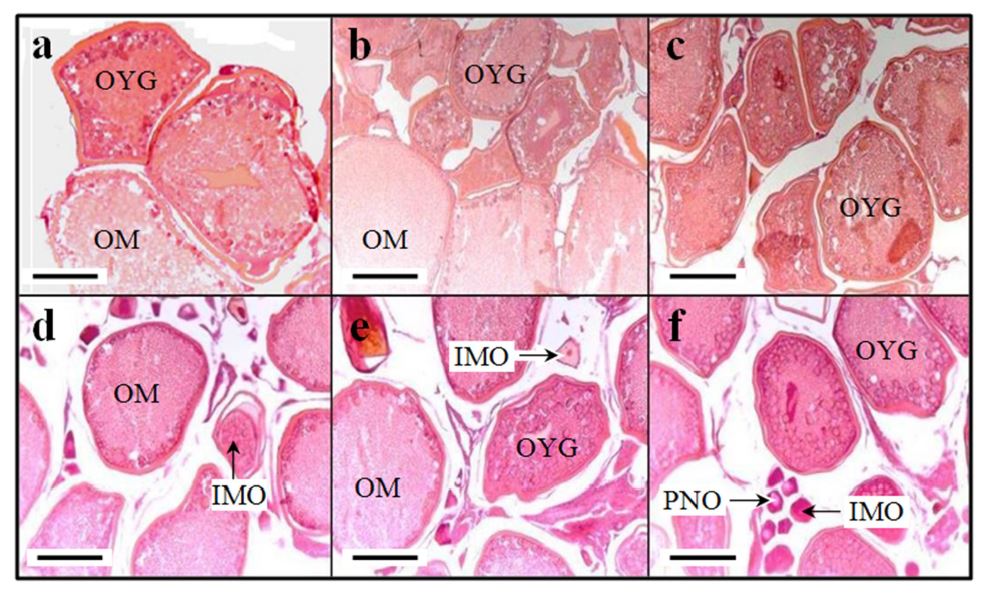
Fig. 5 and 6 shows histological differences of C. auratus, C. cuvieri and common carp between diploid and triploid. Spermatozoon and spermatogonium were shown in testis of diploid C. auratus, C. cuvieri and common carp (Fig. 5- a, b and c). On triploid C. cuvieri and common carp, spermatocyte and spermatogonium were shown in testis, and spermatozoon was not shown in testis (Fig. 5- d and e). Oocytes in yolk granule stage and oocytes in mature stage were observed in ovarium of diploid and triploid C. auratus, C. cuvieri and common carp (Fig. 6- a, b and c). Immature oocytes and oocyte in peri-nucleolus stage were shown in ovarium of triploid C. auratus, C. cuvieri and common carp (Fig. 6- d, e and f). However, Immature oocytes and oocyte in peri-nucleolus stage were not shown in ovarium of diploid C. auratus, C. cuvieri and common carp (Fig. 6- a, b and c). That is, gonads of diploid C. auratus, C. cuvieri and common carp were more mature than those of triploid C. auratus, C. cuvieri and common carp. Gonads of triploid Cyprinidae were not sterility, and were mature slowly than diploid Cyprinidae.

Erythrocyte size, area and volume were significantly different between ploidy and species (Table 4). In all experiment groups, linear length, area and volume of cell were higher than those of nucleus. Linear length of cell and nucleus in triploid species were half as much again as those of diploid in C. auratus, C. cuvieri and common carp (Fig. 7; p<0.05). Area of cell, area of nucleus and volume of cell and nucleus in triploid species were higher than those of diploid species (p<0.05). Linear length of cell and nucleus in diploid C. cuvieri were highest than those in diploid C. auratus and diploid common carp. Also, area of cell, area of nucleus, volume of cell and nucleus in diploid C. cuvieri were highest than those in diploid C. auratus and diploid common carp (p<0.05). Tendency of linear length, area and volume in all triploid species was similar to those of diploid.

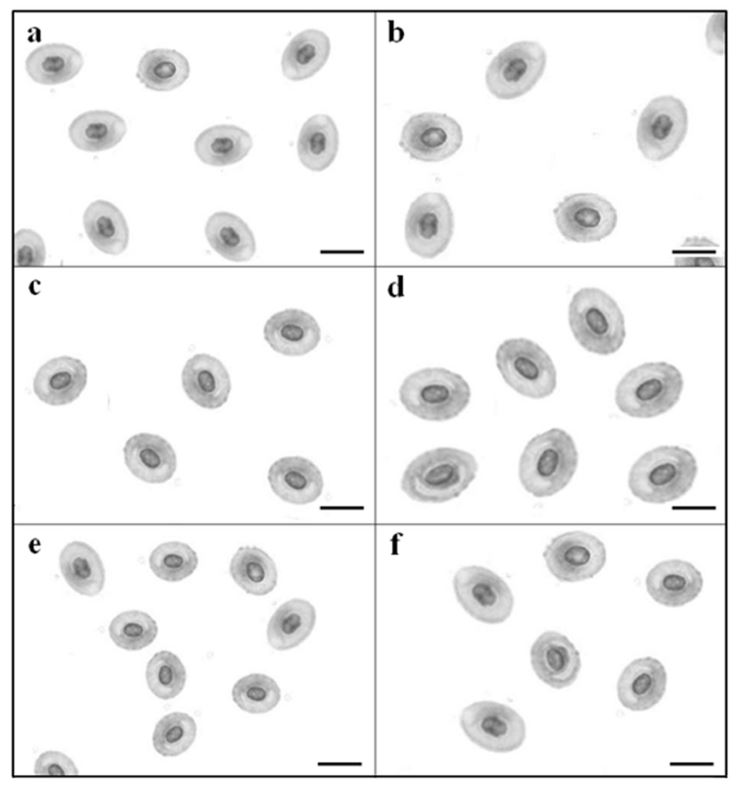
Discussion
In this study, DNA contents of C. auratus, C. cuvieri and common carp were slightly different depending on their habitats. This phenomenon may be due to two reasons. First, the diploid number of chromosomes in C. auratus and common carp is the same, but karyotypes of C. auratus and common carp were different by habitat (Kucinski et al., 2015). For example, C. auratus has 12 metacentrics, 36 submetacentrics and 52 acrocentrics in Japan, and has 24 metacentric, 24 submetacentric and 52 acrocentrics in Romania, Hungary and Ukraine (Kucinski et al., 2015). It means that different karyotype of same species may be caused by a different DNA contents. Next, C. cuvieri has been successfully introduced into the many waters in Korea in the 1970's (Yoon and Park, 2006), and a release of non-native fish into the wild is a serious problem posing considerable ecological and genetic threats through direct competition and hybridization (Kucinski et al., 2015). Rates of hybridization and introgression are increasing worldwide because of translocations of organisms and habitat modifications by humans (Kucinski et al., 2015). It means that used samples in this study may be included hybrid among three species. Therefore, comparative analysis of karyotype in C. auratus, C. cuvieri, common carp and their spontaneous triploid is necessary for detail distinction of C. auratus, C. cuvieri, common carp and their spontaneous triploid in future investigation.
As mentioned by Benfey (1999), triploid cell nuclei contain, by definition, 50% more DNA than diploid cell nuclei. Nuclear volume is increased in triploids to accommodate this extra genetic material. A corresponding increase in cellular volume typically results due to the approximate maintenance of the diploid ratio of nuclear to cytoplasmic volume. Despite increased cell size, triploid individuals are not, as a rule, larger than diploids. This appears to be due to a reduction in cell numbers in those tissues and organs containing larger cells (Benfey, 1999).
In this study, haematological parameters were compared in diploid and triploid specimens of the crucian carp, Carassius auratus, the crucian carp, C. cuvieri and the common carp, Cyprinus carpio. The results showed an increase in erythrocyte size in triploids, in agreement with the previously reported increase in the cell volumes of polyploidy animals (Benfey, 1999). In teleost fish, the increase in erythrocyte size associated with triploidy has already been reported and the measurment of red blood cell dimensions was proposed as a rapid and inexpensive assay for triploidy (Sezaki et al., 1988; Sezaki et al., 1991; Benfey, 1999). Data have usually been obtained from blood cells subjected to air-drying, but this method may lead to alteration in cell morphology. The increase in erythrocyte nuclear size in triploids is a consequence of their higher DNA content (Sezaki et al., 1988; Benfey, 1999).
Spontaneous triploid of C. cuvieri was not reported, while spontaneous triploid of common carp and C. auratus were reported by Al-Sabti et al. (1983), Devlin and Nagahama (2002) and Wu et al. (1993). Females are fertile and males are sterile in triploid hybrids of common carp (Wu et al., 1993). Certain populations of C. auratus can also reproduce by gynogenesis (Kim et al., 2002). For C. auratus, both diploid bisexual and triploid all-female forms have been identified (Boron, 1994; Devlin and Nagahama, 2002). Triploid ova are reported to arise by the formation of a tripolar spindle and abortion of meiosis I, followed by a single nonreductional meiotic division (Cherfas, 1966). Egg activation occurs by fertilization with sympatric Cyprinidae species, but the rare appearance of C. auratus males in gynogenetic populations implies occasional inclusion of paternal genes (Rokicki and Kulikowski, 1994; Devlin and Nagahama, 2002).
In a strain of C. auratus that is unisexual, hormonal masculinization resulted in males with motile but sterile (aneuploid) sperm (Gomels'kii and Cherfas, 1982), as expected from normal meiosis in triploid testis. Allotetraploids (formed between hybrids of C. auratus and C. auratus gibelio or common carp) also can reproduce gynogenetically (Yang et al., 1994), and again, problems with sperm decondensation have been observed. Interestingly, in some populations of gynogenetic C. auratus, both male and female progeny can be found, and both have the same chromosome number within strains (2n = 156, 2n = 166; Shen et al., 1983; Chen et al., 1996), albeit different from that found in other triploid carp gynogenetic fish (2n = 150; Zhou et al., 1983). For gynogenetic triploid C. auratus langsdorfii, formation of triploid ova occurs by suppression of one meiotic division, but recombination still occurs in females, indicating that these gynogenetic fish can reassort sex-determining factors and produce rare male individuals by gynogenetic means (Kobayashi, 1976; Zhang et al., 1992). These examples have revealed how aberrations in chromosome transmission in C. auratus can function to create sex-determination mechanisms that limit the appearance of functional males in populations, although dependence on males from related species for egg activation still remains (Devlin and Nagahama, 2002).
Triploidization is an artificial technique used to generate sterile aquatic animals by taking advantage of the incompatibility in pairing the three homologous chromosomes during meiosis I (Don and Avtalion, 1986). This technique has also been used to enhance the productivity of several fish species because of its assumed ability to increase yield by channeling the energy required from gonadal development to somatic growth (Tave, 1993). More importantly, it generates fish that are unable to breed and contribute to the local gene pool if they were to accidentally escape from confinement. By conferring sterility of exotic fish for a limited purpose, triploidy can serve as an effective method for reducing or eliminating the environmental risks of genetically modified organisms (Murray et al., 1999). Artificial induced triploid fish including Cyprinidae were discovered as sterility by previous reports (Gervai et al., 1980; Seol et al., 2008), but spontaneous triploid Cyprinidae had fertility in our results and previous reports (Al-Sabti et al., 1983; Devlin and Nagahama, 2002; Kim et al., 2002). All measured characteristics of triploid Cyprinidae were half as much again as those of diploid Cyprinidae. We don't determine difference of cell number between diploid and triploid on tissues. Future research will determine cell number of diploid and triploid on tissues, morphometric characteristics on Cyprinidae between diploid and spontaneous triploid, and investigate fertility and spawning detailed mechanism of spontaneous triploid on common carp and C. cuvieri.
- References
-
1. Aliah RS, Inada Y, Yamaoka K, Taniguchi N. 1991. Effects of triploid on haematological characteristics and oxygen consumption of ayu. Nippon Suisan Gakkaishi 57: 833-836.


-
2. Al-Sabti K, Kurelec B, Fijan N. 1983. Spontaneous triploidy and tetraploidy in the common carp (Cyprinus carpio L.). Veterinarski Arhiv 53: 217-223.
-
3. Anjum R, Jankun M. 1994. Spontaneous triploid common carp (Cyprinus carpio L.) in a farm population. Cytobios 78: 153 -157.
-
-
5. Boron A. 1994. Karyotypes of diploid and triploid silver crucian carp Carassius auratus gibelio (Bloch). Cytobios 80: 117-124.
-
6. Chen M, Yang X, Yu X, Chen H. 1996. Karyotype studies on the bisexual natural gynogenetic crucian carp (Carassius auratus) of Pengze. Acta Hydrobiol Sin 20: 25-31.
-
7. Cherfas NB. 1966. Natural triploidy in the females of the unisexual variety of the goldfish Carassius auratus gibelio (Bloch). Sov Genet 13: 557-563.
-
8. Devlin RH, Nagahama Y. 2002. Sex determination and sex differentiation in fish: an overwiew of genetic, physiological, and environmental influences. Aquaculture 208: 191-364.
-
9. Don J, Avtalion RR. 1986. The induction of triploidy in Oreochromis aureus by heat shock. Theor Appl Genet 72: 186-192.
-
-
11. Estevam J, Danaee H, Liu R, Ecsedy J, Trepicchio W, Wyant T. 2011. Validation of a flow cytometry based G2M delay cell cycle assay for use in evaluating the pharmacodynamic response to Aurora A inhibition. J Immuno Meth 363: 135-142.
-
12. Gervai J, Péter S, Nagy A, Horváth L, Csányui V. 1980. Induced triploidy in carp, Cyprinus carpio L. J Fish Biol 17: 667-671.
-
13. Gomel'skii BI, Cherfas NB. 1982. Hormonal reversal of sex in females of a unisexual form of crucian carp. Sov J Dev Biol 13: 142-148.
-
14. Kim EO, Lee JY, Nam YK, Noh JK, Lee SY, Kim DS. 2002. Genetic identification on natural population of triploid crucian carp, Carassius auratus in Korea. J Korean Fish Soc 35: 589-594 (in Korean with English abstract).
-
15. Kim IS. 1997. Illustrated Encyclopedia of Fauna & Flora of Korea, Vol. 37, Freshwater fishes (in Korean). Ministry of Education, Korea, pp 156-509.
-
16. Kim SW, Kim J-M. 2009. Cloning of rod opsin genes isolated from olive flounder, Paralichthys olivaceus, Japanese eel, Anguilla japonica, and common carp, Cyprinus carpio. Fish Aqua Sci 12: 265-275.
-
17. Kobayashi H. 1976. A cytological study on the maturational division in the oogenic process of the triploid ginbuna (Carrassius auratus langsdorfii). Jpn J Ichthyol 22: 234-240.
-
18. Kucinski M, Demska-Zakes K, Zarski D, Liszewski T, Fopp-Bayar D, Jankun M, Furgala-Selezniow G. 2015. The morphological, histological and cytogenetic characteristics of goldfish, Carassius auratus (L.)×common carp, Cyprinus carpio (L.) hybrids. Int J Cytology 68: 77-83.
-
19. Labat R, Hafez R, Quillier E. 1983. Cytogentic studies in some species of Cyprinid fishes. Roczniki Nauk Rolniczych Ser H 150: 2-3.
-
20. Murray JD, Anderson GB, Oberbauer AM, McGloughlin MN. 1999. Transgenic animals in agriculture. In Comparison of traditional breeding and transgenesis in farmed fish with implications for growth enhancement and fitness. Dunham RA, Devlin RH, Eds., CAB International, New York, NY, U.S.A, pp 209-229.
-
21. Nam MM, Yang HJ, Seo BK. 1989. Morphological variation of the crucian carp, Carassius auratus (Linnaeus) from Yongnam area in Korea. Korean J Ichthyol 1: 54-63.
-
22. Rokicki J, Kulikowski M. 1994 Occurrence of male Carassius auratus gibelio (Bloch, 1783) in Poland. Przegl Zool 38: 89-92.
-
23. Seol D-W, Im S-Y, Hur WJ, Park MO, Kim DS, Jo J-Y, Park I-S. 2008. Haematological parameters and respiratory function in diploid and triploid far eastern catfish, Silurus asotus. Gene & Genomics 30: 205-213.
-
24. Sezaki K, Watanabe K, Tsukamoto K, Hashimoto K. 1991. Effects of increase in ploidy status on respiratory function of ginbuna (Carassius auratus langsdrofii) (Cyprinidae). Comp Biochem Physiol A 99: 123-127.
-
25. Sezaki K, Watanabe S, Hashimoto K. 1988. Haematolocical parameters and erythrocyte enzyme activities associated with increase in ploidy status of the spinous loach, Cobitis biwae Jordan and Snyder. J Fish Biol 32: 149-150.
-
26. Shen J, Fan Z, Wang G. 1983. Karyotype studies of male triploid crucian carp (Fangzheng crucian carp) in Heilongjiang. Acta Genet Sin 10: 133-136.
-
27. Tave D. 1993. Growth of triploid and diploid bighead carp (Hypophthalmichthys nobilis). J Appl Aquacult 2: 13-25.
-
28. Wu C, Ye Y, Chen R, Liu X. 1993. An artificial multiple triploid carp and its biological characteristics. Genetics in Aquaculture IV. Aquaculture 111: 255-262.
-
29. Vanparys C, Maras M, Lenjou M, Robbens J, Van Bockstaele D, Blust R, De Coen W. 2006. Flow cytometric cell cycle analysis allows for rapid screening of estrogenicity in MCF-7 breast cancer cells. Toxicol in Vitro 20: 1238-1248.
-
30. Yang ZA, Gui J, Zhu L, Liang S, Jiang Y. 1994. Cytological studies on two differential development modes of the allogynogenetic silver crucian carp multiple tetraploid. Acta Zool Sin 40: 69-74.
-
31. Yoon JM, Park SY. 2006. Genetic comparison between crucian carp (Carassius auratus Linnaeus) and crucian carp (C. cuvieri Termminck and Schlegel). J Anim Sci & Technol 48: 637-650.
-
32. Zhang F, Oshiro T, Takashima F. 1992. Chromosome synapsis and recombination during meiotic division in gynogenetic triploid ginbuna, Carassius auratus langsdorfii. Jpn J Ichthyol 39: 151 -156.
-
33. Zhou J, Shen J, Liu M. 1983. A cytological study on the gynogenesis of Fang-Zheng crucian carp of Heilongjiang Province. Acta Zool Sin 29: 11-15.
Copyright (C) 2025. The Korean Society of Marine Life Science