










JMLS 2016 October;1(2):109-116. Epub 2016 November 22
Copyright © 2016 by The Korean Society of Marine Life Science
Effect of Water Temperature and Photoperiod on the Oxygen Consumption of Four Different Strains of Red Seabream, Pagrus major
Sung-Yong Oh; Marine Ecosystem and Biological Research Center, Korea Institute of Ocean Science & Technology, Ansan 15627, Korea Department of Marine Biology, Korea University of Science and Technology, Daejeon 34113, Korea
Hee Jung Choi; Marine Ecosystem and Biological Research Center, Korea Institute of Ocean Science & Technology, Ansan 15627, Korea
Min-Suk Kim; Marine Ecosystem and Biological Research Center, Korea Institute of Ocean Science & Technology, Ansan 15627, Korea
Yong Joo Park; Marine Ecosystem and Biological Research Center, Korea Institute of Ocean Science & Technology, Ansan 15627, Korea
Jung-Goo Myoung; Marine Ecosystem and Biological Research Center, Korea Institute of Ocean Science & Technology, Ansan 15627, Korea
Joon Yeong Kwon; Department of Aquatic Life Medical Sciences, Sunmoon University, Asan 31460, Korea
Cheol Young Choi; Division of Marine BioScience, Korea Maritime and Ocean University, Busan 49112, Korea
- Abstract
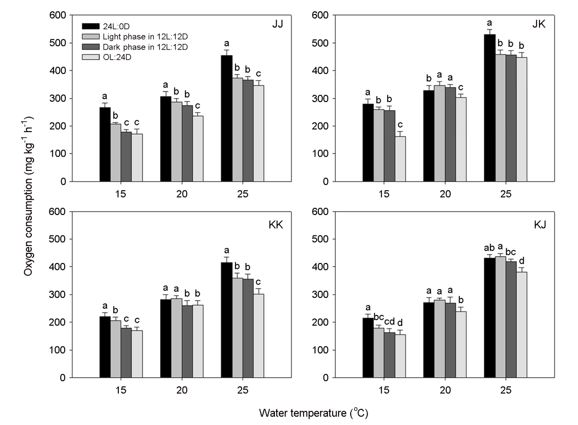
- We determined the effects of different water temperatures (15, 20, and 25℃) and photoperiod cycles (24L:0D, 12L:12D, and 0L:24D) on the oxygen consumption of the offspring of a cultured Japanese strain (JJ), a selected Korean strain (KK), and intraspecific hybrid strains(JK and KJ) of red seabream, Pagrus major, under starvation conditions. The different fish strains, water temperatures, and photoperiod cycles had effects on the mean oxygen consumption of fish. Oxygen consumption increased with increasing water temperaturesfor all photoperiod treatments (p<0.001). Fish held in continuous darkness (0L:24D) used consistently less oxygen than fish exposed to continuous light (p<0.05). The oxygen consumption of fish exposed to the light phase in a 12L:12D photoperiod was higher than that of fish in the dark phase of the 12L:12D cycle, and differences were significant in three of the strains: JJ (15℃), KK (15 and 20℃), and KJ (25℃). The oxygen consumption of the inbred (JJ and KK) and intraspecific hybrid (JK and KJ) strains varied with differing water temperatures and photoperiod cycles. The JK strain displayed significantly higher oxygen consumption than the other strains under all experimental conditions except 15℃ with a 0L:24D photoperiod. The JK and KJ strains usually showed the highest and lowestoxygen consumption values, respectively, whereas the inbred strains exhibited intermediatevalues. Oxygen consumption in the JJ and JK strains was usually higher than that of the KKand KJ strains. We propose that differences in the thermal sensitivity and photosensitization properties of the strains contribute to differences in their ability to adapt to changes in water temperature and photoperiod, thus resulting in differences in the amplitude of their metabolic rates.
Keywords: Pagrus major; Oxygen consumption; Temperature; Photoperiod; Strain
Correspondence to: Sung-Yong Oh Marine Ecosystem and Biological Research Center, Korea Institute of Ocean Science & Technology, Ansan 15627, Korea E-mail : syoh@kiost.ac.kr
- Received
- 10 October 2016;
- Revised
- 17 October 2016;
- Accepted
- 28 October 2016.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: English,
Full Text:

Introduction
Oxygen consumption is a widely used indicator of metabolic rate. Estimates of oxygen consumption rates are essential for assessing the energy metabolism of fish, as well as developing bioenergy models for production (Kaushik, 1998). Oxygen consumption is influenced by numerous internal and external factors, including water temperature (Lyytikäinen and Jobling, 1998), salinity (Forsberg, 1994), fish size (Wuenschel et al., 2004), feed ration (Brett and Groves, 1979), feed composition (Roberts, 1990), photoperiod(Withey and Saunders, 1973), activity (Brett, 1964), and season (Beamish, 1964). Among a variety of environmental factors, water temperature exerts a profound influence on the metabolic reactions that affect all physiological processes in aquatic poikilothermic animals, including fish (Brett and Groves, 1979). Photoperiod is also one of the primary Zeitgebers (i.e, cues or synchronizers) that can influence the daily physiological rhythms of fish (Biswas et al., 2002). Temperature is considered to be a rate-controlling factor, whereas light is classified as a directive factor (Brett, 1979) that stimulates the endocrine system (Björnsson, 1997). The extent to which water temperature and photoperiod combine to influence oxygen con- sumption appears to be specific to distinct fish species (Jonassen et al., 2000; Biswas et al., 2002; Biswas and Takeuchi, 2002). Although the fisheries literature is replete with studies evaluating environ- mental effects, including water temperature and photoperiod, far less research has been conducted on the differences in oxygen consumption between inbred and intraspecific hybrid strains of fish.
The red seabream, Pagrus major, is a commercially important aquaculture fish cultivated in Korea and Japan. Red seabream has been selectively bred for increased growth for the last 20 years in Korea (Noh et al., 2004) and 35 years in Japan (Murata et al., 1996), resulting in the improved growth of this species. Moreover, in Korea, local governments and Korea Institute of Ocean Science & Technology (KIOST) have initiated breeding program protocols aimed to improve growth rates and tentatively establish optimum breeding strains (Noh et al., 2003). Selective breeding programs designed to improve growth rates via better feed utilization have proved successful for several fish species (Gjedrem 1983, 1997; Ogata et al., 2002). However, little information is available on the effects of such selective breeding on the metabolic parameters that affect growth rates, including oxygen consumption. The main objective of this study was to determine the combined effects of water temperature and photoperiod on the oxygen consumption of offspring of a cultured Japanese strain, a selected Korean strain, and intraspecific hybrid strains of red seabream.
3. Statistical analysis
The statistical analysis was performed using MINITAB for Windows (MINITAB Statistical Software, USA, r 13.20). An ANOVA was conducted to identify differences in oxygen consumption among the triplicate experiments and the various treatments. The effects of water temperature on oxygen consumption throughout the experiment were determined by fitting an exponential equation to the data:
where M is the average metabolic rate ($mg$ $O^2$ $kg^{-1}$ $h^{-1}$), T is the water temperature (℃), and a and b are constants.
A one-way ANOVA was used to determine the effects on each strain for all combined treatments. Data were then subjected to Tukey’s test at a 95% significance level to compare the means when differences occurred. Prior to the ANOVA, a Kolmogorov-Smirnov test was used to assess the normality of the distributions, and the homogeneity of variances was assessed using Levene's test.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
***p<0.001
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Strain |
Sources
of |
15℃ |
20℃ |
25℃ |
|||||||||||
|
d.f |
S.S |
M.S |
F |
d.f |
S.S |
M.S |
F |
d.f |
S.S |
M.S |
F |
||||
|
|
Photoperiod |
2 |
120849 |
60424 |
207.01*** |
2 |
60266 |
30133 |
134.60*** |
2 |
155875 |
77937 |
267.58*** |
||
|
JJ |
Levene's
test |
p = 0.614 |
p = 0.253 |
p = 0.237 |
|||||||||||
|
|
Tukey's
test* |
24L:0D>12L:12D>0L:24D |
24L:0D>12L:12D>0L:24D |
24L:0D>12L:12D>0L:24D |
|||||||||||
|
|
Photoperiod |
2 |
187870 |
93935 |
301.65*** |
2 |
18814 |
9407 |
38.37*** |
2 |
97267 |
48633 |
159.06*** |
||
|
JK |
Levene's
test |
p = 0.818 |
p = 0.707 |
p = 0.436 |
|||||||||||
|
|
Tukey's
test* |
24L:0D>12L:12D>0L:24D |
12L:12D>24L:0D>0L:24D |
24L:0D>12L:12D=0L:24D |
|||||||||||
|
|
Photoperiod |
2 |
31177 |
15589 |
73.24*** |
2 |
5096 |
2548 |
8.18** |
2 |
154599 |
77300 |
218.61*** |
||
|
KK |
Levene's
test |
p = 0.302 |
p = 0.534 |
p = 0.994 |
|||||||||||
|
|
Tukey's
test* |
24L:0D>12L:12D>0L:24D |
24L:0D>0L:24D 24L:0D=12L:12D, 12L:12D=0L:24D |
24L:0D>12L:12D>0L:24D |
|||||||||||
|
|
Photoperiod |
2 |
44540 |
22270 |
95.48*** |
2 |
19190 |
9595 |
32.57*** |
2 |
39399 |
19699 |
87.47*** |
||
|
KJ |
Levene's
test |
p = 0.946 |
p = 0.719 |
p = 0.201 |
|||||||||||
|
|
Tukey's
test* |
24L:0D>12L:12D>0L:24D |
24L:0D=12L:12D
>0L:24D |
24L:0D=
12L:12D>0L:24D |
|||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
- References
-
1. Beamish FWH. 1964. Seasonal changes in the standard rate of oxygen consumption of fishes. Can J Zool 42: 189-194.


-
2. Biswas AK, Endo M, Takeuchi T. 2002. Effect of different photo- period cycles on metabolic rate and energy loss of both fed and unfed young tilapia Oreochromis niloticus: Part I. Fish Sci 68: 465-477.
-
3. Biswas AK, Takeuchi T. 2002. Effect of different photoperiod cycles on metabolic rate and energy loss of fed and unfed adult tilapia Oreochromis niloticus: Part II. Fish Sci 68: 543-553.
-
4. Björnsson BT. 1997. The biology of salmon growth hormone: from daylight to dominance. Fish Physiol Biochem 17: 9-14.
-
5. Brett JR. 1964. The respiratory metabolism and swimming perfor- mance of young sockeye salmon. J Fish Res Board Can 21: 1183-1226.
-
6. Brett JR. 1979. Environmental factors and growth. In: Hoar, W.H, Randall, D.J, Brett, J.R (Eds.), Bioenergetics and Growth, Fish Physiology, vol. 8. Academic Press. NY, pp 599-675.
-
7. Brett JR, Groves TDD. 1979. Physiological energetics. In: Hoar WH, Randall DJ, Brett JR (Eds.), Bioenergetics and Growth, Fish Physiology. vol. 8. Academic Press, NY. pp 279-352.
-
8. Chakraborty SC, Ross LG, Ross B. 1992. The effect of photoperiod on the resting metabolism of carp (Cyprinus carpio). Comp Biochem Physiol 101A: 77-82.
-
9. Danzmann RG, Ferguson MM, Allendorf FW. 1987. Heterozygosity and oxygen-consumption rate as predictors of growth and developmental rate in rainbow trout. Physiol Zool 60: 211-220.
-
10. Fonds M, Cronie R, Vethaak AD, Van Der Puly P. 1992. Metab- olism, food consumption and growth of plaice (Pleuronectes platessa) and flounder (Platichthys flesus) in relation to fish size and temperature. Neth J Sea Res 29: 127-143.
-
11. Forsberg OL. 1994. Modeling oxygen consumption rates of post-smolt Atlantic salmon in commercial-scale landbased farms. Aquacul Int 2: 180-196.
-
12. Gjedrem T. 1983. Genetic variation in quantitative traits and selective breeding in fish and shellfish. Aquaculture 33: 51-72.
-
13. Gjedrem T. 1997. Selective breeding to improve aquaculture pro- duction. J World Aquac Soc 28: 33-45.
-
14. Imsland AK, Folkvor A, Stefansson SO. 1995. Growth, oxygen consump tion and activity of juvenile turbot (Scophthalmus maximus L.) reared under different temperatures and photo- periods. Neth J Sea Res 34: 149-159.
-
15. Jonassen TM, Imsland AK, Kadowaki S, Stefansson SO. 2000. In- teraction of temperature and photoperiod on growth of Atlantic halibut Hippoglossus hippoglossus L. Aquac Res 31: 219-227.
-
16. Kaushik SJ. 1998. Nutritional bioenergetics and estimation of waste production in non-salmonids. Aquat Living Resour 11: 211-217.
-
17. Kim IN, Chang YJ, Kwon JY. 1995. The patterns of oxygen con- sumption in six species of marine fish. J Korean Fish Soc 28: 373-381.
-
18. Kim YH, 1999. Oxygen consumption and atmospheric oxygen supply in a recirculating aquaculture system for Nile tilapia culture. Ph.D. Dissertation, Department of Fisheries Biology, Pukyong National University, Busan, Korea, pp 124.
-
19. Lyytikäinen T, Jobling M. 1998. The effect of temperature fluc- tuations on oxygen consumption and ammonia excretion of underyearling Lake Inari Arctic charr. J Fish Biol 52: 1186-1198.
-
20. Mitchell SJ, Cech JJ Jr. 1994. Comparison of metabolic rates of striped bass, white bass, and their hybrid at selected tem- peratures and dissolved oxygen levels. In: MacKinlay DD (Ed.), High Performance Fish. Proceedings of an international fish physiology symposium at the University of British Columbia in Vancouver, 16-21 July 1993, Vancouver, BC, pp 275-279.
-
21. Mitsunaga Y, Sakamoto W, Arai N, Kasai A. 1999. Estimation of the metabolic rate of wild red sea bream Pagrus major in different water temperatures. Bull Japan Soc Sci Fish 65: 48-54.
-
22. Murata O, Harada T, Miyashita S, Izumi K, Maeda S, Kato K, Kumai H. 1996. Selective breeding for growth in red sea bream. Fish Sci 62: 845-849.
-
23. Noh CH, Hong KP, Myoung JG, Kim JM. 2003. Growth perfor- mance of offspring from selected Korean, cultured Japanese and their reciprocal intraspecific hybrids of red seabream, Pagrus major (Abstract). Abstracts and Proceedings of the 2003 Fall Meeting of the Korean Aquaculture Society, 24-25 October 2003, Gangnung, Korea, p 37.
-
24. Noh CH, Hong KP, Oh SY, Choi HJ, Park YJ, Myoung JG, Kim JM. 2004. Comparative growth performance of the selected and the non-selected red seabream (Pagrus major). J Kor Fish Soc 37: 400-404.
-
25. Ogata HY, Oku H, Murai T. 2002. Growth performance and macronutrient retention of offspring from wild and selected red seabream (Pagrus major). Aquaculture 206: 279-287.
-
26. Oh SY, Jang YS, Park HS, Choi YU, Kim CK. 2012. The influence of water temperature and body weight on metabolic rate of olive flounder Paralichthys olivaceus. Ocean Polar Res 34: 93 -99.
-
27. Oh SY, Noh CH. 2006. Effects of water temperature and photo- period on the oxygen consumption rate of juvenile dark-banded rockfish, Sebastes inermis. J Aquaculture 19, 210-215.
-
28. Peck MA, Buckley LJ, Bengtson DA. 2004. Inter-individual differ- ences in rates of routine energy loss and growth in young-of-the-year juvenile Atlantic cod. J Fish Biol 64: 984-995.
-
-
30. Ross LG, McKinney RW. 1988. Respiratory cycles in Oreochromis niloticus (L.) measured using a six-channel microcomputer-operated respirometer. Comp Biochem Physiol 89: 637-643.
-
31. Withey KG, Saunders RL. 1973. Effect of reciprocal photoperiod regime on standard rate of oxygen consumption of postsmolt Atlantic salmon (Salmo salar). Can J Fish Aquat Sci 30: 1898 -1900.
-
32. Wuenschel MJ, Werner RG, Hoss DE. 2004. Effect of body size, temperature and salinity on the routine metabolism of larval and juvenile spotted seatrout. J Fish Biol 64: 1088-1102.
Copyright (C) 2024. The Korean Society of Marine Life Science