










JMLS 2019 December;4(2):53-62. http://jmls.or.kr Epub 2019 December 13
Copyright © 2019 by The Korean Society of Marine Life Science
Bacterial Distribution and Relationship with Phytoplankton in the Youngsan River Estuary
Se Hee Kim; Department of Ocean System Engineering, Mokpo National Maritime University, Mokpo 58628, Korea
Yong Sik Sin; Department of Ocean System Engineering, Mokpo National Maritime University, Mokpo 58628, Korea
- Abstract
해양 종속영양 박테리아는 수생태계에서 미소생물환의 주요 구성원으로 유기물의 분해 등 생지화학적 순환에서 중요한 역할을 한다. 해양 생태계의 물질 순환과 에너지 흐름을 이해하기 위해서는 박테리아의 변동에 대한 연구 조사가 중요하다. 본 연구는 방조제 건설로 변형된 영산강 하구 해수역을 대상으로 박테리아와 환경인자들의 월 변동 양상을 조사하였고, 박테리아 변동의 주요인자인 식물플랑크톤(chlorophyll-a)과의 상관성을 크기별로 구분하여 파악하고자 하였다. 연구 결과, 영산강 하구의 박테리아는 저층보다 표층에서 높은 개체수를 보였으며, 겨울철보다 여름철에 개체수가 높았다. 그리고 방조제에 가까울수록 개체수가 증가하였으며, 방조제와 가장 인접한 정점에서 2018년 8월, 9월 그리고 2019년 6월에 최대치로 증가하였다. 박테리아의 개체수가 높았던 정점과 시기에 식물플랑크톤의 생체량도 증가하면서 통계분석결과에서도 양의 상관성을 보였고 크기별로도 모두 유사한 상관성을 보였다. 이러한 결과는 식물플랑크톤 기원의 유기물이 박테리아 변동에 영향을 미치고 있고 크기별로 그 영향의 차이가 없음을 제시하고 있다. 또한 수온에 비례하여 증가하는 박테리아의 계절 분포는 박테리아의 성장에 대한 수온의 영향을 보여주는 결과라 볼 수 있다. 그 외에 간헐적인 담수 유입을 통한 영양염 공급과 박테리아의 개체수 변동의 연관성은 관찰되지 않았다. 본 연구에서는 또한 특정시기에 성층이 거의 없는 조건에서 용존산소가 고갈되는 빈산소층이 관찰되었는데 이는 식물플랑크톤 기원 유기물 공급과 박테리아의 분해로 인한 산소 소모의 결과로 추정된다.
Heterotrophic bacteria are a major member of the microbial loop in the marine ecosystem and play an important role in the biogeochemical cycle decomposing organic matter. Therefore study of bacterial variation is important to understand the material cycle and energy flow of marine ecosystems. We investigated the monthly variations of bacteria and environmental factors in the Youngsan River estuary, and the correlation between bacteria and phytoplankton biomass (chlorophyll-a) including size-structure. As a result, bacteria of the Youngsan River estuary were higher in the surface than in the bottom layer, and higher in the summer than in winter. And the closer to the dike, the abundance increased, and it increased to the peaks in August, September, and June 2019 at the station closest to the dike. The chlorophyll-a also increases at the stations and time when the bacterial abundance was high and they correlates positively displaying no difference between size fractions. The results indicate that organic matter derived from phytoplankton has an effect on bacterial variation but no size-dependent effects. In addition, the seasonal pattern of bacteria increasing in proportion to the water temperature suggests the effect of water temperature on the growth of bacteria. No association of bacterial abundance variation with nutrient supply due to freshwater input was observed. In this study, dissolved oxygen was depleted and hypoxia was observed for a short time when a strong stratification was not developed. This may be resulted from the supply of organic matter from phytoplankton and the consumption of oxygen due to bacterial decomposition.
Keywords: Bacteria(박테리아) Youngsan River estuary(영산강 하구) Monthly variation (월 변동) Phytoplankton(식물플랑크톤) Size-structure(크기구조)
Correspondence to: Yong Sik Sin; Department of Ocean System Engineering, Mokpo National Maritime University, Mokpo 58628, Korea; E-mail : yongsik@mmu.ac.kr;
- Received
- 23 November 2019;
- Revised
- 24 November 2019;
- Accepted
- 9 December 2019.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
해양 종속영양 박테리아는 미소생물환(Microbial loop)의 주요 구성원으로서(Azam et al., 1983; Sherr and Sherr, 1987; Pomeroy and Wiebe, 1993) 유기물 분해와 영양염 재생산 및 흡수 등을 통해 해양의 생지화학적 순환에 매우 중요한 역할을 담당한다(Cole et al., 1988; Ducklow and Carlson, 1992). 따라서 해양 생태계의 물질 순환과 에너지 흐름을 이해하기 위해서는 박테리아와 같은 이차생산자의 변동에 대한 연구 조사가 중요하다(Sim et al., 2010). 또한, 박테리아는 용존유기물(DOM)을 분해하는 과정에서 산소를 소모하기 때문에 수층 내 저산소 및 무산소 환경 형성에 직접적인 영향을 미칠 수 있다(McDonough et al., 1986; Cole and Pace, 1995; Ochs et al., 1995; Rabalais et al., 2002). 결국 해양생태계를 이해하고 관리하기 위해서는 박테리아의 변동 조사를 포함한 생태 연구가 필요한 상황이다.
미소생물환에서 종속영양 박테리아의 탄소원인 용존유기물의 주요 공급원은 식물플랑크톤으로(Azam et al., 1983) 이들 미생물간의 상호작용은 수질이나 생태계 건강성에 영향을 미칠 수 있다. 특히, 육지로부터 공급되는 영양염으로 인해 매우 생산적인 시스템으로 보고되어 있는(Ryther, 1969) 하구에서는 부영양화 여건에서 식물플랑크톤 대증식이 발생할 수 있기 때문에 이들의 상호작용에 대한 이해가 수질 및 생태계 관리 측면에서 필수적이다. 하구는 담수와 해수가 혼합되는 전이수역으로 조석, 파랑, 담수 유입 등과 같은 수리역학적 요인의 영향을 받아 물리 · 화학적, 생물학적으로 복잡한 환경이다. 따라서 하구의 박테리아 분포와 식물플랑크톤의 상호작용에 대한 조사는 해양미소생물환의 구조를 파악하는데 중요한 정보를 제공할 수 있다.
영산강 하구는 1981년에 방조제가 축조되면서 자연형 하구에서 인공형 하구로 변형되었다. 방조제 내측은 담수역, 외측은 해수역으로 분리되었으며, 담수역에서는 생활하수 및 농 · 축산 폐수의 유입으로 영양염 농도가 증가할 수 있는 여건을 가지고 있다(Cho and Park, 1998). 더불어 2011년에는 4대강 사업의 일환으로 승촌보와 죽산보가 건설되었고, 이로 인해 수체의 흐름이 정체되면서 녹조가 빈발하는 등 수질 악화의 원인이 될 것으로 예상되고 있다(Sin et al., 2015). 이러한 담수가 간헐적으로 방조제 수문을 통해 해수역으로 유입되면서 하구의 수질 및 식물플랑크톤 생체량에 크게 영향을 주는 것으로 보고되었다(Yoon et al., 2013; Sin and Jeong, 2015). 이렇게 인위적으로 담수가 유입되는 변형 하구에서 박테리아가 반응하는 형태 그리고 식물플랑크톤과의 상호작용에 대한 연구(Kim, 2015)는 매우 미흡한 상황이다. 특히 식물플랑크톤을 크기별로 구분하여 박테리아와 상관성을 조사한 연구는 전무하다. 식물플랑크톤 크기구조는 수생태계 먹이망을 통한 에너지 흐름을 결정한다고 보고된 바 있다(Walsh, 1976). 따라서 본 연구에서는 영산강 하구 해수역을 대상으로 박테리아와 식물플랑크톤의 월 변동과 기타 환경인자들을 조사하여 박테리아 변동에 영향을 미치는 주요인자를 파악하고자 하였다. 특히 박테리아 변동과 식물플랑크톤 크기구조와의 상관성은 식물플랑크톤 생체량을 크기별로 구분(size-fractionated chlorophyll-a)하여 조사하였다.
Ra = Fluorometer 측정값($\rm {\mu g ~ l^{-1}}$)
결 과
1. 물리 · 화학적 환경인자
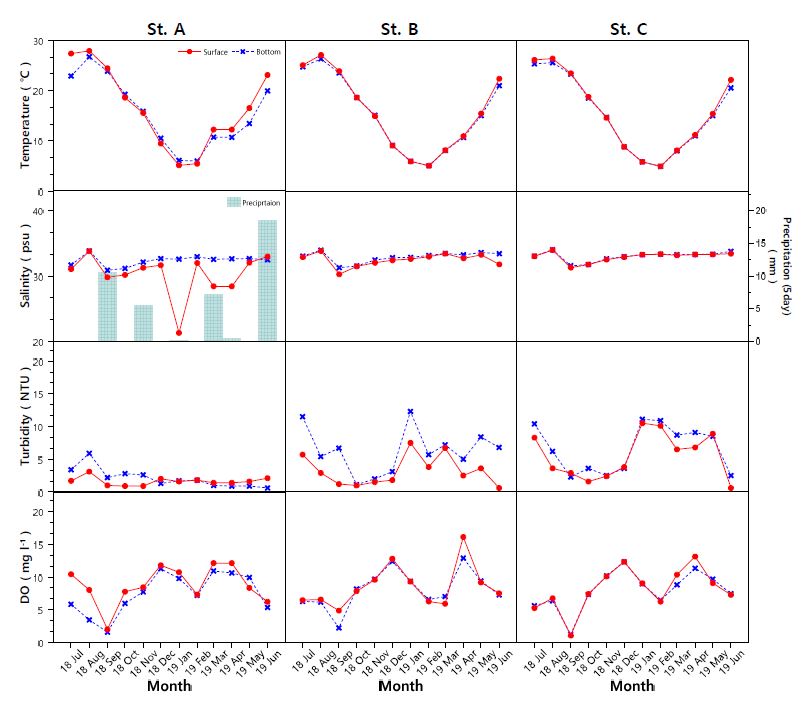
수온의 분포를 보면 4.9~27.9℃로 겨울철에 낮고, 여름철에 높은 계절적인 경향을 나타냈다(Fig. 2). 수문을 통해 방류되는 담수의 영향을 직접적으로 받을 수 있는 A 정점은 조사기간 동안 대부분의 시기에 표 · 저층간 수온차(-0.3~4.5℃)를 보였으며, 특히 2018년 10월에서 2019년 2월까지는 수온 역전(-1.1~-0.3℃) 현상이 발생하였다. 담수 유입의 영향을 직접적으로 판단할 수 있는 지표인 염분은 21.3~34.0 psu 분포를 보였으며 방조제 인근인 A 정점에서 외해인 C 정점으로 갈수록 표 · 저층의 염분차가 감소하였다(Fig. 2). A 정점은 일부 시기(2018년 8월, 2019년 6월)를 제외하고 조사기간 동안 저층의 염분 농도가 표층보다 높게 나타났다. 5일 동안의 누적 강수량은 0.2~18.5 mm의 분포를 보였다. 강수량이 없었던 2018년 1월에는 표층 염분이 21.3 psu까지 감소하였는데 이는 방조제를 통한 인위적 담수 유입의 영향을 보여주는 결과라 할 수 있다. 탁도의 표 · 저층 분포는 A 정점에서 0.6~5.9 (1.9±1.1) NTU, B 정점 0.6~12.3 (4.8±3.2) NTU, C 정점 1.6~11.1 (6.1±3.5) NTU로, 세 정점 간에 서로 다른 농도 분포를 보이며 외해인 C 정점으로 갈수록 높은 분포를 나타냈다(Fig. 2). 또한 A 정점에서는 2018년 7~11월에 2.2~5.9 NTU로, B, C 정점은 0.6~12.3 NTU로 대부분의 시기 저층에서 더 높은 탁도 분포를 보였다. 용존산소는 세 정점의 표 · 저층에서 각각 1.58~12.14 $\rm {mg ~ l^{-1}}$, 2.20~16.11 $\rm {mg ~ l^{-1}}$, 1.04~13.12 $\rm {mg ~ l^{-1}}$의 분포를 보였다(Fig. 2). 2018년 9월 시기에 A, C 정점에서 각각 1.6~2.0 $\rm {mg ~ l^{-1}}$, 1.0~1.2 $\rm {mg ~ l^{-1}}$로 빈산소층이 형성되었으며 B 정점은 2.2~4.9 $\rm {mg ~ l^{-1}}$로 빈산소에 가까운 농도로 감소하였다.
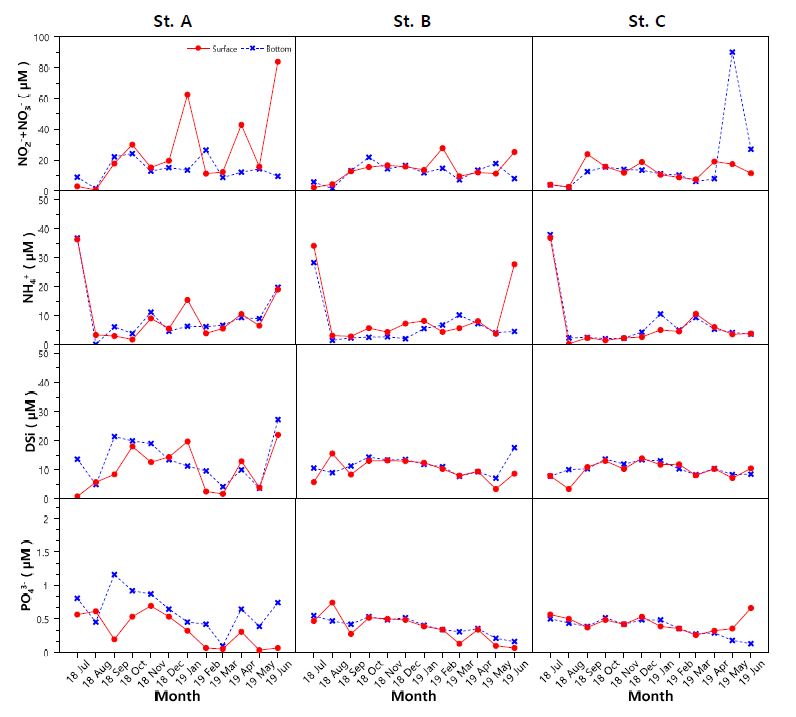
영양염의 분포를 살펴보면(Fig. 3), 아질산+질산성 질소($\rm NO_2^-$ + $\rm NO_3^-$)는 0.57~90.08 μM의 분포를 보였으며, 2019년 1월, 4월, 6월에 A 정점에서 표층 농도가 42.83~83.87 μM로 높게 나타냈다. 같은 시기에 표층 규산염(DSi) 또한 12.85~21.97 μM로 높은 분포를 보였다. 암모니아성 질소($\rm NH_4^+$)는 0.14~37.90 μM의 분포를 보였으며, 2018년 7월에 28.27~37.90 μM로 세 정점에서 모두 높은 분포를 나타냈다. 인산염($\rm PO_4^{3-}$)은 0.03~1.16 μM의 분포를 보였다.
2. 크기별 chlorophyll-a
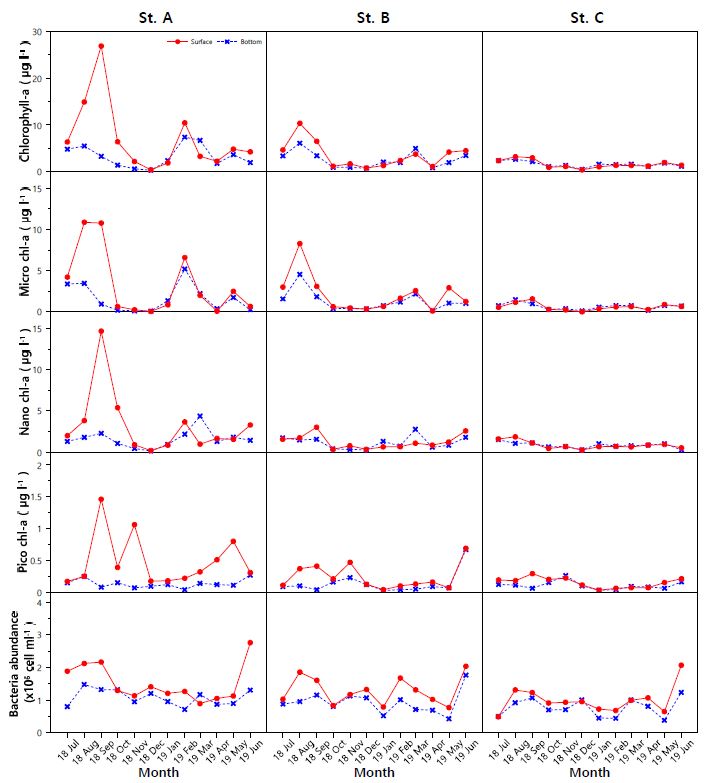
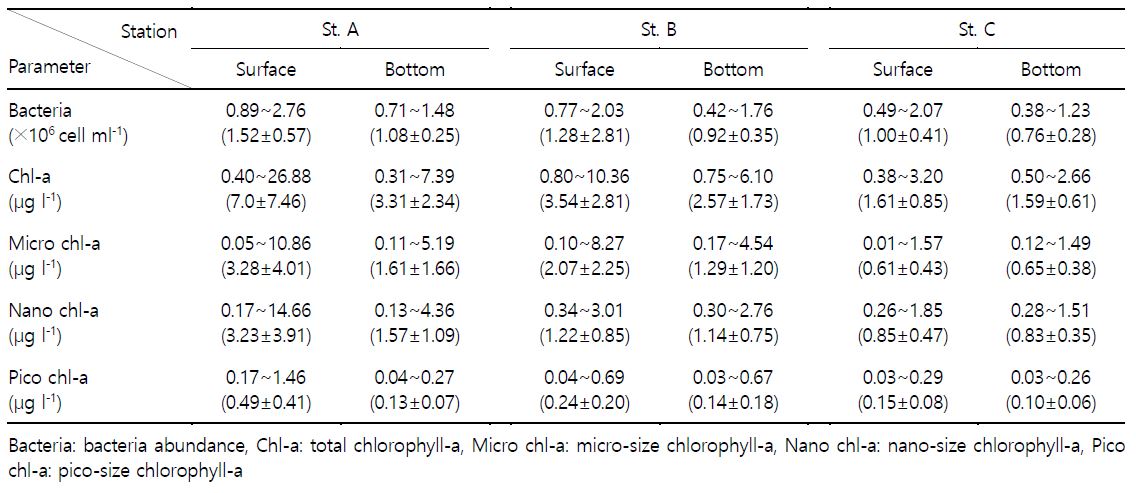
식물플랑크톤 생체량의 지표인 chlorophyll-a는 세 정점의 표 · 저층에서 0.31~26.88 $\rm {\mu g ~ l^{-1}}$의 분포를 나타냈다(Fig. 4). 시간적으로는 2018년 8, 9월에 2.17~26.88 (7.33±7.20) $\rm {\mu g ~ l^{-1}}$로 높은 표 · 저층 분포를 보였으며 12월에 0.94~6.40 (0.52±0.20) $\rm {\mu g ~ l^{-1}}$로 낮은 표 · 저층 분포를 나타냈다. A 정점의 표 · 저층 분포는 2019년 2월에 7.39~10.45 $\rm {\mu g ~ l^{-1}}$로 다시 증가하는 경향을 보였고, B 정점은 3월에 3.76~4.98 $\rm {\mu g ~ l^{-1}}$, C 정점은 5월에 1.85~1.97 $\rm {\mu g ~ l^{-1}}$의 농도로 다소 증가하는 경향을 나타냈다. Chlorophyll-a는 방조제 인근 정점일수록 높은 농도를 보였으며(Table 1), 크기별 chlorophyll-a 또한 같은 양상을 나타냈다. 크기별 chlorophyll-a 월 변동을 살펴보면(Fig. 4), 2018년 8, 9월에 micro-size가 1.16~10.86 $\rm {\mu g ~ l^{-1}}$로 높은 분포를 보였다. 또한 9월에는 nano-size가 1.11~14.66 $\rm {\mu g ~ l^{-1}}$로 높은 분포를 나타냈다. 2018년 11, 12월에 micro-size는 0.01~0.46 $\rm {\mu g ~ l^{-1}}$로 낮은 분포를 나타냈으며 nano-size 또한 12월에 0.13~0.33 $\rm {\mu g ~ l^{-1}}$로 낮은 분포를 보였다. 전체적으로 높은 chlorophyll-a 분포를 보였던 9월에 pico-size는 표층에서 0.29~1.46 $\rm {\mu g ~ l^{-1}}$로 높게 나타났으나 저층은 0.04~0.08 $\rm {\mu g ~ l^{-1}}$로 낮은 분포를 보였다.
3. 박테리아 개체수
영산강 하구에서 2018년 7월부터 2019년 6월까지 박테리아 변동을 살펴보면(Fig. 4), 표층은 0.49~2.76 (1.27±0.51)×$10^6$ cell $\rm {ml^{-1}}$의 분포를 보였으며 저층은 0.38~1.48 (0.92±0.32)×$10^6$ cell $\rm {ml^{-1}}$로 나타나 전체적으로 표층의 개체수가 높았다. 표 · 저층 평균 농도는 A~C 정점에서 각각 1.30×$10^6$ cell $\rm {ml^{-1}}$, 1.10×$10^6$ cell $\rm {ml^{-1}}$, 0.88×$10^6$ cell $\rm {ml^{-1}}$로 나타났고 방조제에 가까운 A 정점으로 갈수록 증가하는 경향을 보였다(Table 1). 시간적으로는, 2018년 8, 9월에 세 정점의 표층에서 1.23~2.16×$10^6$ cell $\rm {ml^{-1}}$의 높은 분포를 보였으며, 9월에는 저층에서도 1.06~1.32×$10^6$ cell $\rm {ml^{-1}}$로 높은 개체수를 보였다. 또한, 12월에 저층은 1.00~1.20×$10^6$ cell $\rm {ml^{-1}}$로 높은 분포를 보였다. 2019년 5월은 세 정점의 표 · 저층에서 0.38~1.12×$10^6$cell $\rm {ml^{-1}}$로 모두 낮은 분포를 보였다가, 6월에 1.23~2.76×$10^6$ cell $\rm {ml^{-1}}$로 다시 증가하여 높은 분포를 나타냈다.
4. 상관분석
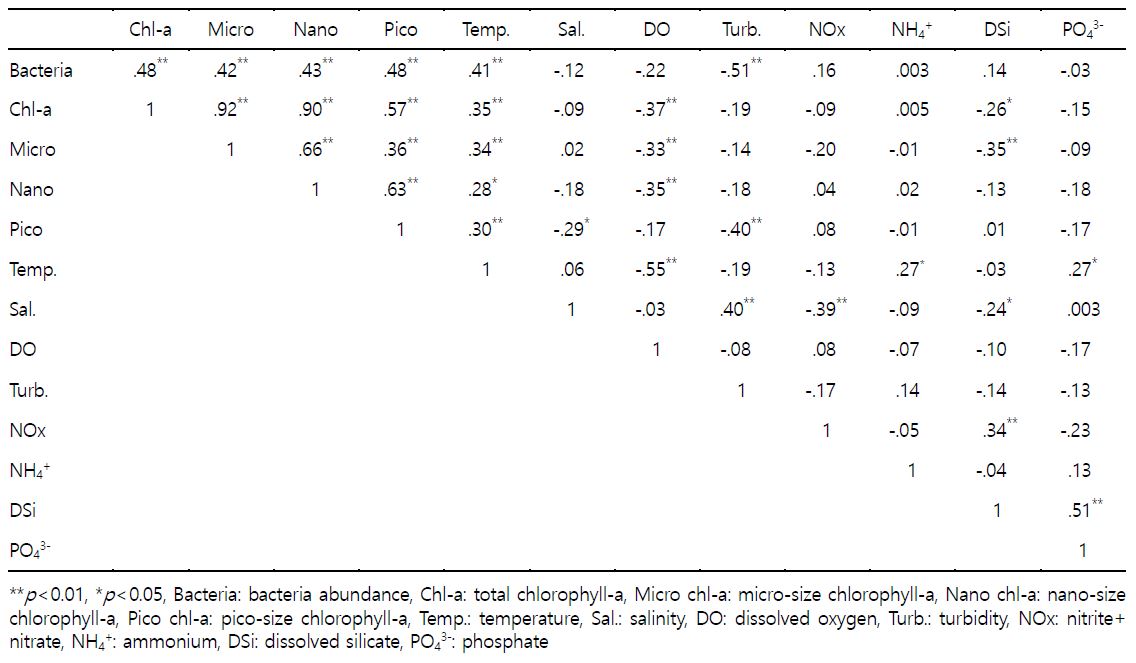
박테리아와 크기별 chlorophyll-a, 물리 · 화학적 환경인자들과의 관계를 파악하기 위해 상관성을 분석한 결과(Table 2), 박테리아 개체수는 크기별 chlorophyll-a (micro-, nano-, pico-size), 수온과 유의한(p<0.01) 양의 상관관계를 보인 반면 탁도와는 유의한(p< 0.01) 음의 상관성을 보였다. 크기별 chlorophyll-a는 수온과 양의 상관관계를 보였다. 또한 micro-, nano-size는 용존산소와 유의한 음의 상관관계를 나타냈다. Pico-size는 염분, 탁도와 음의 상관관계를 나타내 다른 크기(micro-, nano-)와 다른 양상을 보였다. 수온은 용존산소와 유의한(p<0.01) 음의 상관관계를 나타냈으며, 암모니아성 질소, 인산염과 유의한(p<0.05) 양의 상관관계를 보였다. 염분은 탁도와 유의한(p<0.01) 양의 상관관계를 나타냈으며, 아질산+질산성 질소, 규산염과 유의한(p<0.01, p<0.05) 음의 관계를 보였다.
- References
-
1. Azam F, Fenchel T, Field JG, Gray JS, Meyer-Rei LA, Thingstad F. 1983. The ecological role of water column microbes in the sea. Mar Ecol Progr Ser 126: 97-102.


-
2. Bird DF, Kalff J. 1984. Empirical relationships between bacterial abundance and chlorophyll concentration in fresh and marine waters. Can J Fish Aquat Sci 41: 1015-1023.
-
-
4. Cho BC, Choi JK, Chung CS, Hong GH. 1994. Uncoupling of bacteria and phytoplankton during a spring diatom bloom in the mouth of the Yellow Sea. Mar Ecol Prog Ser 115: 181 -190.
-
5. Cho BC, Park MG, Shim JH, Choi DH. 2001. Sea-surface temperature and f-ratio explain large variability in the ratio of bacterial production to primary production in the Yellow Sea. Mar Ecol Progr Ser 216: 31-41.
-
6. Cho BC, Shim JH. 1992. Significance of Estuarine Mixing in Distribution of Bacterial Abundance and Production in the Estuarine System of the Mankyung River and Dongjin River, Korea. J Kor Soc Oceanogr 27: 154-163.
-
7. Cho YG, Park KY. 1998. Heavy metals in surface sediments of the Youngsan Estuary, West Coast of Korea. J Environ Sci Int 7: 549-557.
-
8. Cole JJ, Findlay S, Pace ML. 1988. Bacterial production in fresh and saltwater ecosystem: a cross-system overview. Mar Ecol Pro Ser 43: 1-10.
-
9. Cole JJ, Pace ML. 1995. Bacterial secondary production in oxic and anoxic freshwaters. Limnol Oceanogr 40: 1019-1027.
-
10. Diaz RJ, Rosenberg R. 1995. Marine benthic hypoxia: A review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanogr Mar Biol Ann Rev 33: 245-303.
-
-
12. Ducklow HW, Carlson CA. 1992. Oceanic bacterial production: In Advances in microbial ecology. Springer, Boston, MA 12: 113 -181.
-
13. Felip M, Pace ML, Cole JJ. 1996. Regulation of planktonic bacterial growth rates: The effects of temperature and resources. Micro Ecol 31: 15-28.
-
14. Findlay S, Pace ML, Lints D, Howe K. 1992. Bacterial metabolism of organic carbon in the tidal freshwater Hudson estuary. Mar Ecol Prog Ser 89: 147-153.
-
15. Hoch MP, Kirchman DL. 1993. Seasonal and inter-annual variability in bacterial production and biomass in a temperate estuary. Mar Ecol Prog Ser 98: 283-295.
-
16. Jardillier L, Basset M, Domaizon I, Belan A, Amblard C, Richardot M, Debroas. 2004. Bottom-up and top-down control of bacterial community composition in the euphotic zone of a reservoir. Aquat Microb Ecol 35: 259-273.
-
17. Kim SH. 2015. Ecological and biogeochemical processes in the Yeongsan River estuary, Yellow Sea. Department of environmental marine sciences. Doctor of philosophy. Graduate school of Hanyang University.
-
18. Kimura H, Izumi M. 1994. Effect of oxygen deficiency of the sea bottom on environment of bay. Fish Eng 31: 41-45.
-
19. Kormas KA, Kapiris K, Thessalou-Legaki M, Nicolaidou A. 1998. Quantitative relationships between phytoplankton, bacter and protists in an Aegean semi-enclosed embayment (Maliakos Gulf, Greece). Aquat Microb Ecol 15: 255-264.
-
20. Lee CW, Kudo I, Yanada M, Maita Y. 2001. Bacterial abundance and production and heterotrophic nanoflagellate abundance in subartic coastal waters (Western North Pacific Ocean). Aquat Microb Ecol 23: 263-271.
-
21. McDonough RJ, Sanders RW, Porter KG, Kirchman DL. 1986. Depth distribution of bacterial production in a stratified lake with an anoxic hypolimnion. Appl Environ Microbiol 52: 992-1000.
-
22. Naganuma T, Miura S. 1997. Abundance, production and viability of bacterioplankton in the Seto lsland Sea. Japan J Oceanogr 53: 435-442.
-
23. Ochs CA, Cole JJ, Likens GE. 1995. Population dynamics of bacterioplankton in an oligotrophic lake. J Plankton Res 17: 365 -391.
-
24. Painchaud J, Therriault JC. 1989. Relationships between bacteria, phytoplankton and particulate organic carbon in the Upper St. Lawrence Estuary. Mar Ecol Prog Ser 56: 301-311.
-
25. Parsons TR, Maita Y, Lalli C. 1984. A manual of chemical and biological method for seawater analysis. Pergamon Press Inc., New York, pp 173.
-
26. Plummer DH, Owens NJP, Herbert RA. 1987. Bacteria—particle interactions in turbid estuarine environments. Cont Shelf Res 7: 1429-1433.
-
27. Pomeroy LR, Wiebe WJ. 1993. Energy sources for microbial food webs. Mar Microb Food webs 7: 101-118.
-
28. Porter KG, Feig YS. 1980. The use of DAPI for identifying and counting aquatic microflora. Limnol Oceanogr 25: 943-948.
-
29. Rabalais NN, Turner RE, Wiseman Jr WJ. 2002. Gulf of Mexico hypoxia, aka "The dead zone". Ann Rev Ecol Evol S 33: 235 -263.
-
30. Ritter C, Montagna PA. 1999. Seasonal hypoxia and models of benthic response in a Texas bay. Estuaries 22: 7-20.
-
-
32. Sherr EB, Sherr BF. 1987. High rates of consumption of bacteria by pelagic ciliates. Nature 325: 710-711.
-
33. Shiah FK, Ducklow HW. 1994a. Temperature regulation of heterotrophic bacterioplankton abundance, production, and specific growth rate in Chesapeake Bay. Limnol Oceanogr 39: 1243 -1258.
-
34. Shiah FK, Ducklow HW. 1994b. Temperature and substrate regulation of bacterial abundance, production and specific growth rate in Chesapeake Bay, USA. Mar Ecol Prog Ser 103: 297-308.
-
35. Shim JH, Shin YK, Cho BC. 1993. The roles and the inter relationship between bacteria and phytoplankton in estuarine system of Mankyung and Dongjin rivers. Korea J of Kor Soc Oceanogr 28: 107-113.
-
36. Sim JH, Kim YS, No I, Park YC, Park JG, Park C, Sin YG, Yang SR, Yang JS, Yeo HG, Yu SJ, Lee DS, Lee WH, Jeong HJ, Jo BC, Cha SS, Choi JK. 2010. Heterotrophic micro-plankton. Plankton ecology. 4st ed. Seoul National University, Korea. pp 155-171.
-
37. Simon M, Cho BC, Azam F. 1992. Significance of bacterial biomass in lakes and the ocean: comparison to phytoplankton biomass and biogeochemical implications. Mar Ecol Progr Ser 86: 103 -110.
-
38. Sin Y, Jeong B. 2015. Short-term variations of phytoplankton communities in response to anthropogenic stressors in a highly altered temperate estuary. Estuar Coast Shelf Sci 156: 83-91.
-
39. Sin Y, Lee E, Lee Y, Shin K-H. 2015. The river estuarine continuum of nutrients and phytoplankton communities in an estuary physically divided by a sea dike. Estuar Coast Shelf Sci 163: 279-289.
-
40. Stachowitsch M. 1984. Mass mortality in the Gulf of Trieste: the course of community destruction. Mar Ecol 5: 243-264.
-
41. Vaquer-Sunyer R, Duarte CM. 2008. Thresholds of hypoxia for marine biodiversity. Proceedings of the National Academy of Sciences of the United States of America 105: 15452-15457.
-
42. Wainright SC. 1990. Sediment-to-water fluxes of particulate material and microbes by resuspension and their contribution to the planktonic food web. Mar Ecol Prog Ser 62: 271-281.
-
43. Walsh JJ. 1976. Herbivory as a factor in patterns of nutrient utili- zation in the sea. Limnol Oceanogr 21: 1-13.
-
44. Weiss RF. 1970. The solubility of nitrogen, oxygen and argon in water and seawater. Deep Sea Res 17: 721-735.
-
45. Weisse T. 1991. The annual cycle of heterotrophic freshwater nomoflagellates: role of bottm-up versus top-down control. J Plankton Res 13: 167-185.
-
46. White PA, Kalff J, Rasmussen JB, Casol JM. 1991. The effect of temperature and algal biomass on bacterial production and specific growth rate in freshwater and marine habitats. Microb Ecol 21: 99-118.
-
47. Yang EJ, Choi JK, Hyun JH. 2003. The study on the seasonal variation of microbial community in Kyeonggi Bay, Korea. I. Bacteria and heterotrophic nanoflagellates. J Kor Soc Oceanogr 8: 44-57.
-
48. Yoon B-B, Lee E-J, Kang T-A, Shin Y-S. 2013. Long-term change of phytoplankton biomass (chlorophyll-a), environmental factors and freshwater discharge in Youngsan Estuary. Korean J Ecol Environ 46: 205-214.
-
49. Zarkanellas AJ. 1979. The effects of pollution-induced oxygen deficiency on the benthos in Elefsis Bay, Greece. Mar Environ Res 2: 191-207.
Copyright (C) 2024. The Korean Society of Marine Life Science